8.5. Первичные аттрактанты. V - защита и место для выведения потомства
В литературе часто встречается понятие Obdachblume - "цветок-убежище". К этому понятию очень часто прибегают, когда не находят каких-либо других причин для объяснения того или иного явления. Несомненно, что шмели часто остаются на ночь и ищут убежища в цветках (самцы это делают постоянно), однако очень сомнительно, чтобы это играло какую- либо существенную роль для процесса опыления, даже если и наблюдаются определенные преимущества (Peponapis в семействе Cucurbitaceae). Фогель сообщал нам, что он находил насекомых, прятавшихся в цветках Serapias, но ван дер Пэйлу никогда не удавалось находить насекомых в цветках этого рода в дневное время. Горовиц (Horovitz, 1976) утверждает, что у Anemone coronaria "наиболее существенными векторами пыльцы служат те насекомые, которые используют цветки в качестве убежища и двигаются беспорядочно от цветков в мужской стадии... к цветкам в женской стадии... в меньшей степени пчелы, такие, как самцы Eucera sp. и ряд других - от питающихся пыльцой жуков до хищных представителей семейства Anthocoridae".
Достаточно убедительный пример использования цветка как убежища при изменении температуры - это использование цветка Calluna vulgaris. Очевидно, что цветки Calluna приспособлены к посещению крупными насекомыми, и пчелы охотно его посещают, но эти визиты могут быть излишними из-за активности Taeniothrips ericae (Hagerup, 1950а; Haslerud, 1974), которые проводят большую часть своей жизни внутри цветка.
Животные, которые не очень активно оберегают свое потомство, откладывают яйца там, где есть пища, например в цветке. И если Calluna представляет собой сочетание как убежища, так и места для выведения потомства, то Trollius europaeus служит примером последнего. Опыление в этом случае еще весьма "грязное". Более тонкий процесс, представляющий продолжение этого пути развития, обнаружен Райли у Yucca и юкковой тегитикулы. Позднее это явление тщательно исследовалось другими авторами (Baker, 1961b; Powell, Mackie, 1966). Иногда утверждают, что те адаптации, которые встречаются в экологии опыления, не являются взаимными; растения якобы приспосабливаются к (статическим) животным так же, как они приспосабливаются к ветру или воде как опыляющим агентам. Нет лучшего опровержения этому, чем пример Tegiticula ("Pronuba"), этологию и морфологию которой можно понять только в том случае, если рассматривать их как часть общего синдрома опыления ("межвидовой симбиоз"; Алпатов).
Другими примерами совершенной взаимной адаптации между цветком и насекомыми, ищущими место для выведения потомства, служат известные случаи Ficus spp., характеризующиеся очень сложной сменой поколений как у растения, так и у опылителя. Более простое отношение описано Брухом (Bruch, 1932) для Prosopanche burmeisteri (Hydnoraceae). В толстых мясистых цветках этого покрытосеменного паразитического растения обычно выводятся личинки опыляющих их жуков; эти личинки питаются только наружными частями цветков и поэтому не мешают дальнейшему развитию растения. Интересно в этом отношении близкое сходство между цветками Prosopanche и стробилами Cycadeoidea, причем последний опыляется таким же путем, что было показано Крепетом (Crepet, 1972).
Ни в какой другой области экологии опыления равновесие не является столь неустойчивым, как в синдроме места для выведения потомства, а именно равновесие между взаимной пользой и деструктивным поведением по отношению к опылителю или цветку. Первый может съесть семена, а второй - индуцировать "ложную кладку яиц".
Паразитическая роль некоторых видов ночных бабочек (Dianthoecia, Hadena), опыляющих определенные виды Caryophyllaceae, а также Lycaena spp. в других семействах, известна уже давно (Kerner, 1898). При этом наблюдается в основном деструктивный эффект - личинки поедают развивающиеся семязачатки. Однако деструкция не всегда носит общий характер: некоторые коробочки остаются нетронутыми (либо потому, что в них не были отложены яйца, либо потому, что яйца погибли), и в них развиваются семена. В этом случае опыление необходимо и для насекомого, иначе не было бы пищи для развивающихся внутри гинецея личинок. По данным Стиртона (Stirton, 1976), другой представитель Hadeniae паразитирует на семенах, но в то же самое время является единственным опылителем некоторых Liliaceae в Южной Африке.
Во взаимоотношениях между Hadena bicruris и Melandrium album, как показал Брантез (Brantjes, 1976), весьма существенны следующие факторы:
- самки должны питаться перед каждой кладкой яиц;
- и самки, и самцы питаются как на тычиночных, так и на пестичных цветках;
- яйца откладываются (в естественных условиях) только на пестичных цветках.
Если каждый цветок дает некий объем нектара (или другой пищи), равный v, а ночной бабочке перед откладыванием яиц необходимо получить V нектара, то она должна посетить V/v цветков. Если считать, что самцы делают то же самое, то число посещений увеличивается вдвое, однако только половина насекомых вернется к женскому цветку. Если в каждый период яйцекладки будет отложено е яиц и каждая личинка уничтожит с пестиков из общего числа С, приходящегося на 1 цветок, то мы получим:
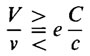
Если в каждый период яйцекладки будет отложено е яиц и каждая личинка уничтожит с пестиков из общего числа С, приходящегося на 1 цветок, то мы получим
При равенстве этого выражения требования насекомого удовлетворены полностью, но растение не получает ничего. Если левая часть неравенства меньше правой, то насекомое будет ощущать недостаток в пище, и некоторые личинки погибнут от голода; только в том случае, если левая часть неравенства будет больше правой, станет возможным опыление растения. В случае Yucca v, V е и С равны единице, и "излишек для опыления" получается вследствие того, что с меньше единицы (личинки не съедают весь пестик). У Trollius (С/с) > 1 и опять наблюдается эффект опыления. У Melandrium album результат опыления, по-видимому, очень незначителен и, возможно, опыление и завязывание семян обусловлено активностью других насекомых. В данном случае стратегия опыления в сообществе как едином целом противопоставляется стратегиям отдельных особей. Эту проблему, которая постоянно возникает как одна из основных на последних дискуссиях, мы обсудим ниже. В этой связи отношения Hadena - Melandrium можно рассматривать как тип паразитического поведения, которое становится таковым в силу определенных обстоятельств и поэтому не может существовать без давления извне; должен существовать какой-то альтернативный процесс, в противном случае взаимоотношения прекратятся. Могут ли такие взаимоотношения развиваться дальше вплоть до достижения коадаптации, подобной коадаптации у Yucca, не ясно, и мы этого касаться не будем.
Трудно не рассматривать подобные синдромы опыления как стадии в экологической эволюции, которая могла иметь место и в различных группах насекомых и во многих группах растений, и которая шла от более или менее случайных, более или менее дистропных отношений к совершенному циклическому симбиозу, когда цветок и опылитель следуют в своем развитии друг за другом и обоюдно взаимозависимы.
Готтсбергер (Gottsberger, 1970) обнаружил, что жуки-опылители (долгоносики), выводящие свое потомство в цветках Annonaсеае, делают это несколькими способами: одни личинки превращаются в куколку внутри цветоножки, другие живут в опадающих очень толстых лепестках. Однако поскольку цветки представляют собой необычный субстрат для этих личинок, отношения не носят характера собственно симбиоза, хотя они прекрасно служат целям растения.
Описано (Silberbauer-Gottsberger, 1973; Essig, 1971, 1973) опыление, которое производят во время кормления и размножения дистропные жуки в еще закрытых обертках различных родов пальм. Аналогичное наблюдение было сделано и относительно масличной пальмы (Heusser, 1912). У Salacca обнаружена двойная система привлечения: пчел (Trigona) привлекает (ночью) нектар, и в то же самое время жуки, откладывающие яйца, посещают соцветия (Mogea, 1978).
Подобный тип аттрактации (места для выведения потомства) не зависит от организации цветка покрытосеменных, как это наблюдается в идентичном случае для стробилов саговниковых (Wester, 1910). По данным Готтсбергера (Gottsberger, 1970), виды, выводящие потомство в стробилах Zamia, обнаруживались во взрослом состоянии внутри цветков Annonaceae. Специфичность аттрактации, типичная для обычного синдрома опыления, по-видимому, не развивается.
Не очень близкие взаимоотношения наблюдаются между мельчайшими двукрылыми (Forcipomya sp.) и растением какао. Насекомое выводит свое потомство в разлагающихся бобах, и это наводит на мысль о том, что слишком безжалостная чистка плантаций может помешать получить семена в следующем году (Dessart, 1961). Однако Африка, где это описано, не является родиной Theobroma, и, возможно, в Америке эти взаимоотношения совсем иные. Сходная ситуация была описана Уиндером и Силвой (Winder, Silva, 1972 a, b), причем в этом случае местом выведения потомства служили представители семейства бромелиевых.
Сходная ситуация описана Шнейдером и Муром (Schneider, Moore, 1977) для жуков Donacia piscatrix и Nymphaeaceae, которым присущ синдром опыления, характерный для всего семейства. Копуляция обычно происходит внутри или около цветков (аттрактация - свидание), яйца откладываются под листьями, личинки проникают внутрь и кормятся около растущей зоны корневища. Будучи деструктивным, это поведение предполагает существование равновесия между растением и опылителем-хищником.
Особым случаем аттрактации, относящейся к синдрому убежище - место выведения потомства, является использование цветков хищниками, скрывающимися в них. Горовиц и др. (Horowitz et al. 1975) описали случай потенциального опыления Anemone coronaria хищными клопами в Израиле. Этот синдром характерен также для арктических областей (Kevan, 1972).
При обманной аттрактации (в редких случаях) процесс может зайти так далеко, что насекомое откладывает яйца в цветке, но они, как правило, не имеют возможности развиваться, хотя в некоторых цветках они все же развиваются, образуя новое поколение опылителей. Так обеспечивается пополнение опылителей. То же самое наблюдается у Artocarpus heterophyllus, относящегося к другому анемофильному роду (van der Pijl, 1953), близкому к Ficus. Борг (Borg, 1977) обнаружил личинок внутри тычиночных соцветий других Моrасеае: Cecropia, Trilapsium, что делает переход более правдоподобным. Положение Dorstenia в этом ряду не ясно. Сходные примеры имеются в других семействах: Alocasia pubera (van der Pijl, 1933), Rafflesia и Philodendron, опыляемый жуками (Hubbard, 1895). В цветки Colocasia откладывают яйца мухи (Cleghorn, 1913), а в Dieffenbacha - жуки (Madison, 1979). В большинстве случаев эти синдромы можно рассматривать как синдромы, развившиеся из более простого сапромиофильного синдрома.
В этой связи мы снова обращаемся к наблюдению Рэтрея (Rattray, 1913) над жуками, поедающими пыльцу из мужских шишек Encephalartos и откладывающих яйца в женских. Очевидно, два типа шишек воздействуют на различные инстинкты насекомого - единственный известный до сих пор случай. Однако что же привлекает насекомых, откладывающих яйца в мужские цветки других двудомных растений? Для человека мужские шишки Encephalartos имеют запах, а женские - нет.
В первых изданиях этой книги мы упоминали цветки орхидеи Oberonia twaithessii, по внешнему виду напоминающие тлей, и предположили, что они могут привлекать муравьев. Так как это было только предположение, а дополнительных данных по этому вопросу не поступало, мы решили опустить ссылку. Однако в 1977 г. Иври и Дафни показали, что Epipactis consimilis помимо нектарной аттрактации и аттрактации места совокупления привлекает также женские особи мух-журчалок, личинки которых питаются растительными тлями. Самки откладывают яйца на растение; это, как полагают авторы, возможно потому, что цветки по своей окраске, размеру (и запаху) имитируют тлей. На наш взгляд, цветки Oberonia еще больше напоминают тлей.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'