5.2. Структура и функция цветка покрытосеменных
Части простейшего цветка, представленного на рис. 1, имеют очень четкие функции, которые можно проследить без каких-либо ссылок на "адаптацию". Так, обычная функция чашечки - защита развивающегося цветка до опыления. У некоторых растений эта функция настолько ярко выражена, что чашечка опадает или засыхает в начале цветения (например, у мака). Функция венчика состоит в привлечении опылителей. В пыльниках образуется пыльца, а в пестике располагается семязачаток. Рыльце представляет собой воспринимающую поверхность, а столбик несет рыльце и обеспечивает рост пыльцевой трубки к семязачатку.
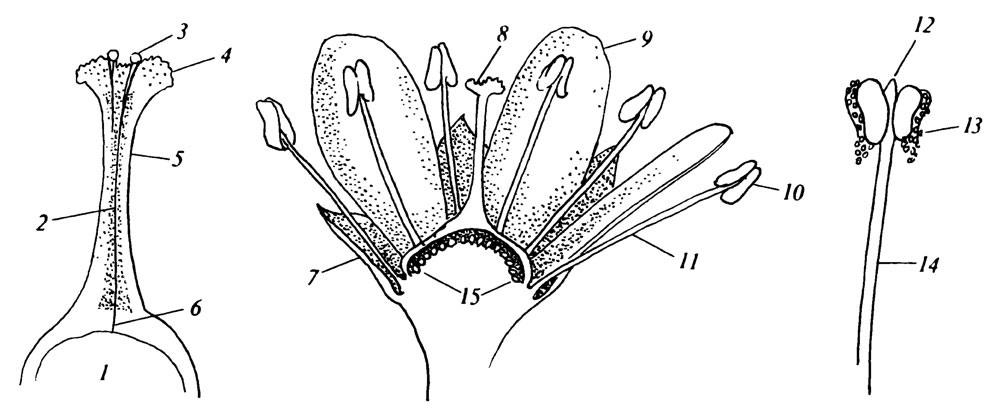
Рис. 1. Основные части цветка. 1 - завязь; 2 - проводниковая ткань; 3 - пыльцевые зерна; 4 - рыльце; 5 - столбик; 6 - пыльцевая трубка; 7 - чашелистик; 8 - гинецей (пестик); 9 - лепесток венчика; 10 - андроцей; 11 - тычинка; 12 - связник; 13 - пыльник; 14 - тычиночная нить; 15 - семязачатки
Это краткое описание позволяет связать определенные морфологические единицы с их наиболее общей функцией, но бывает так, что у некоторых цветков отдельные функции отсутствуют или осуществляются другими органами. В этом случае орган морфологически отличается от своего "обычного состояния; в нем возникают "адаптации" к новой функции. Так, защитную функцию могут выполнять брактеи (Compositae) или какие-либо другие выросты чашечки, а также и сегменты венчика (Umbelliferae). Для привлечения насекомых могут служить не только венчик, но и чашечка (Calluna), тычинки (Саnnа) или пестик (Iris), или каждый в отдельности, или в сочетании друг с другом; для этой же цели могут служить и нецветковые части растения (Cornus).
Образование пыльцы происходит только в пыльниках, но преподношение ее насекомым могут осуществить и другие органы. Все эти функциональные модификации становятся более понятными, если рассматривать их в связи со специфическим способом опыления - они образуют часть синдрома опыления. Мы не будем описывать морфологию наружных частей цветка, поскольку таких описаний очень много; строение же тычинок и пыльцы требует более подробного рассмотрения*. У очень примитивных цветков, например у Degeneria (А. С. Smith, 1949), тычинка сходна с (микро) спорофиллом криптогамного или саговникового, с теками, расположенными как отдельные спорангии на небольшом спорофилле (или погруженными в него). У более развитых в эволюционном отношении цветков ламинарная часть тычинки сильно редуцирована, но тека еще сохраняет характер спорангия и имеет специальный механизм открывания. Раскрывание спорангия начинается с появления щели вдоль теки; при этом стенка так изгибается, что выворачивает теку наружу. Пыльца насекомоопыляемых растений прилипает к внешней поверхности пыльника, пока ее не удалит опылитель.
* (Мы не будем касаться спорных вопросов о морфологической природе "спорофилла" и происхождения спорангия. Для обсуждения современных теорий о морфологической природе и происхождении цветка покрытосеменных мы отсылаем читателей к статьям Готтсбергера (Cottsberger, 1974) и Мёзе (Meeuse, 1975), с которыми мы, однако, не всегда согласны. Мы не поддерживаем мнения о том, что деление на спорофиллы и семафиллы, проведенное Леппиком (Leppik, 1961), полезно для экологии опыления; также не согласны с высказыванием Mëze (Meeuse, 1974), что "все ламиниформные элементы функционируют как оптические аттрактанты в репродуктивной сфере зоофильных растений". Оно не объясняет функцию привлечения, характерную для спорофиллов, которая очень важна для некоторых цветков, а также не принимает во внимание привлечение запахом, которое в примитивных цветках играет даже большую роль, чем оптическое привлечение. Возможно, что первые "семафиллы" выполняли скорее функцию удержания, чем функцию привлечения.)
Сухая пыльца ветроопыляемых растений обычно удаляется из пыльника довольно быстро. Однако из этих двух правил бывают исключения.
Пыльники могут раскрываться с помощью крышечек или клапана, как у Berberidaceae, или могут выделять пыльцу через поры (Solanaceae, Cassia); иногда они вытягиваются в трубку, как у Ericaceae, а иногда опылитель удаляет часть стенки теки, как, например, у некоторых видов Garcinia. Многие из этих необычных механизмов открывания являются частью синдрома специфического опыления. В открывающихся порами пыльниках, например у Rhododendron, Exacum и у многих Melastomataceae, пыльца выжимается или выбрасывается (van der Pijl, 1939). В пыльниках некоторых цветков имеются специальные придатки (рожки), которые, двигаясь, ударяют по пыльнику, вызывая тем самым высыпание пыльцы (Vaccinium, Thunbergia, см. гл. 17). У многих Mimosoideae на вершине пыльника находится желёзка, функция которой пока не ясна.
У Salvia, например S. pratensis (см. разд. 13.2), стерильная средняя часть пыльника - связник - очень длинная (вернее, очень широкая) и соединяется с короткой жесткой тычиночной нитью; кроме того, в этом случае одна тека стерильна. Эта сложная структура, которая с определенными модификациями обнаружена также у Calceolaria (Vogel, 1974) и Roscoea (Nordhagen, 1932), выполняет определенную функцию при посещении цветков насекомыми. И в других семействах связник бывает очень сильно развит, например в семействе Melastomataceae. У Canna большая часть тычинок стерильна и является вместе с околоцветником органом привлечения; только одна тычинка фертильная, причем она в свою очередь имеет только одну фертильную теку, остальные - лепестковидные.
Вторичное преподношение пыльцы (не путать со случайными нарушениями) наблюдается в тех случаях, когда пыльца высыпается из теки, обычно перед или в начале цветения, и откладывается в другой части цветка, причем это сопровождается завяданием или более или менее полным исчезновением пыльника. У Proteaceae пыльца попадает в ту часть цветка, которая развивается позже в зрелое рыльце. Campanula (см. разд. 17.1) и многие представители Compositae, отдельные цветки которых функционируют, подобно цветкам Campanula, представляют классический пример этого. У родственного семейства Goodeniaceae морфологическое развитие пошло еще дальше, например в роде Scaevola на вершине столбика расположена чашевидная структура (плюска). Перед цветением пыльца собирается в плюске, а затем выталкивается рыльцем (Kugler, 1973). Еще более развитый механизм обнаружен у некоторых растений, например у видов Centaurea. У этих растений пыльца не выделяется до тех пор, пока опылитель не начнет раздражать тычиночные нити. Только тогда начинает выделяться пыльца. Вторичное преподношение пыльцы с использованием частей пестика наблюдается не только в родственных Campanulales, но и в других семействах, например Rubiaceae, цветки которых имеют тенденцию к двудомности; при этом столбик служит для преподношения пыльцы у тычиночных цветков. Подобная же структура обнаружена в соцветиях различных Compositae, у которых отдельные цветки утратили пестичную функцию, но тем не менее имеют пестик как орган для преподношения пыльцы (van der Pijl, 1978; Stuessy, 1972).
Более общий случай представляет механизм у Centaurea, функция которого заключается в защите пыльцы, причем выполняется она в этом случае очень скрытно. Пыльца нуждается в защите от насекомых, не принимающих участие в опылении, от сдувания ветром (за исключением анемофильных растений), от вымывания во время дождя, и прежде всего от влаги, которая может убить пыльцу некоторых видов (см. Kerner, 1873) или, например, вызвать преждевременное ее прорастание (Eisikowitch, Woodell, 1975).
Очень простым и эффективным способом защиты от намокания является поникшее положение многих цветков; правда, это положение имеет и другие эффекты (см. Hallermeier, 1922). Закрывание всего цветка или каких-либо отдельных его частей также обеспечивает хорошую защиту от воды (см. цветки Pedicularis, обсуждение разд. 17.8). В других случаях пыльца укрыта в узких трубочках, в которые вода проникает с трудом, или же защищена покровом из волосков или чешуек, также предотвращающих попадание воды в цветок. Следует отметить, что волоски в цветках не всегда выполняют эту функцию, и, кроме того, имеются пыльцевые зерна, на которые вода не оказывает никакого вредного действия, по крайней мере в короткие промежутки времени. Многие цветки с наступлением плохой погоды закрываются. Такой способ имеет большое преимущество, поскольку пыльца и нектар (если он имеется) оказываются укрытыми на тот период, пока не функционируют опыляющие агенты (ветер или животное). Например, периодическое раскрывание цветков (дневных и ночных), вероятно, имеет одинаковое значение как способ защиты пыльцы и нектара в те периоды, когда нет опыляющих агентов. Пример с Centaurea, описанный выше, представляет еще один шаг в этом направлении: пыльца освобождается только в тот момент, когда опылитель садится на этот цветок.
По-видимому, не все движения тычинок носят приспособительный характер. Однако необходимо учитывать, что многие структуры цветка ранее считались нефункциональными, и только при более тщательном исследовании было установлено, что они занимают важное место в синдроме опыления; именно поэтому мы не хотим утверждать категорически, что какие-то движения тычинок не несут вовсе никакой функции, даже если некоторые случаи сейчас объяснить нельзя.
После созревания пыльца может жить в течение более или менее длительного времени. Обычно продолжительность ее жизни составляет несколько дней, но известны как более короткие, так и более длительные периоды (см. Visser, 1955; Brewbaker, 1959; Pruzcinsky, 1960). Пыльца, собранная в корзиночки перепончатокрылых, сразу же теряет способность к прорастанию (Maurizio, 1959). Это происходит под действием жирной (10-окси-2-деценоевой) кислоты, вырабатываемой челюстными железами насекомых (Lokoschus, Keulart, 1968). В экспериментальных условиях продолжительность жизни пыльцы может быть значительно увеличена. По данным Фешнера и Фунша (Fechner, Funsch, 1966), пыльца Pinus ponderosa прорастала через И лет после хранения при низкой температуре (0-4 С) и средней влажности (25-30%). Хэнсон и Кэмпбелл (Hanson, Campbell, 1972) сообщили о таких же результатах при хранении после лиофильной сушки, а Коллинз и др. (Collins et al., 1973) получили положительные результаты при хранении пыльцы в жидком азоте.
В естественных условиях продолжительность жизни пыльцевых зерен зависит от скорости их метаболизма; метаболически более активные трехъядерные зерна живут обычно недолго. По данным Тикменева (Tikhmenev, 1974), жизнеспособность пыльцевых зерен арктических злаков измеряется, вероятно, несколькими часами. У Digitalis пыльца может терять способность к прорастанию уже к концу цветения (Daumann, 1970 b). Вместе с тем в Ботаническом саду Копенгагена удалось произвести оплодотворение у Cycas circinalis пыльцой, которая хранилась при обычной температуре в течение 11 мес. (Sørensen 1970). Следует отметить также чрезвычайно медленное развитие процесса оплодотворения у многих цветущих ранней весной деревьев, например Corylus. Пыльцевое зерно начинает прорастать сразу же, но затем молодая пыльцевая трубка длительное время остается в покое, после чего возобновляет дальнейший рост.
Влажная пыльца часто оказывается неспособной к прорастанию. Это может быть следствием выхода из экзины легко растворимых белков (ферментов) (Knox, Heslop-Harrison, 1971), хотя роль этих белков в процессе прорастания, если она и существует, пока еще не доказана.
Гинецей является более консервативной структурой, чем наружные части цветка, и по форме напоминает бутылку. В некоторых семействах (Orchidaceae, Asclepiadaceae) верхние части гинецея и андроцея срастаются, образуя одну центральную структуру - колонку, в которой тека образует карманы.
Наиболее изменчивой частью гинецея является рыльце. Обычно оно образует небольшую липкую выпуклость или щель на верхушке столбика; у Iris и Sarracenia активная поверхность рыльца составляет очень незначительную часть от всей его поверхности. Размер функционально активной части рыльца можно определить с помощью химических реакций, например реакцией с перекисью водорода, предложенной Зейслером (Zeisler, 1938). Столбик и рыльце у представителей некоторых родов развиваются довольно необычно, хотя значения этих отклонений не всегда ясны. Прилипание пыльцевых зерен к рыльцу обусловлено частично их собственной способностью, частично клейкой поверхностью рыльца, а частично его шероховатостью. Относительное значение каждого из этих факторов меняется у различных растений. Рыльце иногда расположено в цветке так, что оно счесывает пыльцу со щеточек на задних ножках опылителей, как, например, у Iris.
Функция рыльца, как уже было сказано, состоит в том, что оно является воспринимающей поверхностью для пыльцевых зерен. Специфические требования для прорастания пыльцевых зерен различны: некоторые из них легко прорастают на искусственной среде, для других необходимы добавки определенных химических веществ, а третьи совсем не могут прорастать на искусственной среде, как, например, пыльцевые зерна Pavetta javanica, которые прорастают только на рыльцах, того же вида и P. fulgens. Взаимодействие рыльца и пыльцевого зерна будут обсуждаться в разд. 5.3.
При изучении цветка покрытосеменных основной проблемой является покрытосемянность. Как показывают геологические данные, покрытосемянность дает растениям большие преимущества, и покрытосеменные с момента своего появления доминируют и превосходят численно все другие группы. Конечно, причины могут быть и в других признаках, сопутствующих покрытосемянности (Fægri, 1974), но абсолютно точно можно говорить о том, что покрытосемянность не является невыгодным условием. Термин "покрытосемянность" не совсем точен: укрыты не семена, а семязачатки. Поэтому привычное название "покрытосемянность" лучше бы заменить на "покрытосемязачатковость".
Некоторые из результатов, обусловленных покрытосемянностью, легко себе представить. Отрицательная сторона заключается в том, что процесс опыления и оплодотворения стал более сложным. С этим связано образование целого ряда новых органов: рыльца, столбика, проводниковой ткани в женской сфере и длинных пыльцевых трубок в мужском гаметофите. Это, между прочим, изменило и функцию гаметофита: от ризоидоподобного питающего органа (не очень отличающегося от микрогаметофита, например, у Laminaria) до очень специализированного органа для переноса мужского ядра. Имеются данные, правда не очень убедительные, о том, что и у хвойных, например у Tsuga, наблюдается тенденция в направлении покрытосемянности (см. Doyle, 1945). Отсутствие у некоторых покрытосеменных, например у Degeneria, типичных рылец может быть примитивным свойством, но может быть и следствием вторичного развития (например, у Tambourissa(?) ср. Endress, 1979, 1980).
Положительное свойство покрытосемянности - надежная защита семязачатка от высыхания. Тот факт, что у наземных растений, например у Taxus, существуют полностью незащищенные семязачатки, не опровергает сам по себе полезность этой функции, особенно при продвижении растений в засушливые районы; следует, однако, указать, что внешняя механическая защита от высыхания в большей степени может быть заменена внутренним физиологическим механизмом. В связи с покрытосемянностью следует заметить, что процесс оплодотворения у покрытосеменных в меньшей степени зависит от водной среды, чем у любой другой группы растений, за исключением высших мхов. Таким образом, вряд ли можно объяснить всеобщий успех покрытосемянности только воздействием ее на условия влажности в гинецее.
Второй вопрос, который нам следует рассмотреть, заключается в том, что поверхность рыльца и ткани столбика проявляют избирательность в отношении прорастания пыльцевого зерна и роста пыльцевой трубки. Это будет обсуждаться в разд. 5.3. Избирательность проявляется в задержке прорастания несовместимых пыльцевых зерен и создании тем самым больших возможностей для перекрестного опыления - более выгодного способа опыления у высших растений (см. Whitehouse, 1950). Если бы не было такой избирательности ("сита"), единственной преградой к самоопылению была бы несовместимость между геномами при предполагаемом слиянии ядер. Таким образом, эффект "сита" имеет наибольшее селективное значение и может, несомненно, способствовать успеху покрытосемянности. Весьма сомнительно, однако, что приведенных доводов достаточно для объяснения современного превосходства покрытосеменных.
Третье значение покрытосемянности состоит в защите семязачатка от механических воздействий. Это становится особенно важным, если считать, что предки покрытосеменных опылялись животными, возможно, неуклюжими питающимися пыльцой жуками (см. разд. 11.1.1). Надежное укрытие семязачатков для защиты их от грубых прикосновений, возможно, имело в этом случае большие селективные преимущества, как утверждали некоторые ботаники, подойдя к проблеме с различных сторон. Эта точка зрения позволила рассмотреть в более широком биологическом смысле перигинию и эпигинию:

Третье значение покрытосемянности состоит в защите семязачатка от механических воздействий
Происхождение покрытосеменных вряд ли когда-нибудь удастся доказать с помощью палеонтологических данных, и для решения этого вопроса должны быть предложены гипотезы. Однако если рассматривать цветок как функциональную единицу в отношении опыления, то все его особенности, включая покрытосемянность, будут представлять логичную систему. Тот факт, что морфологические элементы, обусловливающие эпигинию и перигинию сильно различаются в различных семействах (Douglas, 1957), может служить указанием на то, что они возникли в соответствии с функциональными требованиями, а не как некоторый вид морфологического ортогенеза (см. V Grant, 1950 а)*. Для выяснения вопроса о происхождении покрытосемянности, возможно, имеет значение обнаружение на микроспорах (или "плодах") Caytoniales - одной из групп растений, независимо достигших покрытосемянности, фекалий крошечных рептилий, живущих на них.
* (Поскольку несомненно, что птицы-опылители также касаются цветка весьма грубо и что в этом случае очень выгодна дополнительная защита, мы не решаемся принять идею Гранта о том, что опыление птицами является очень древним и предшествует опылению шмелями. Вместе с тем очевидно, что более высокоорганизованные опылители - перепончатокрылые и чешуекрылые - касаются цветков очень осторожно, поэтому против них достаточно даже самой слабой защиты.)
Пестик имеет две основные функции: 1) защита семязачатков от неблагоприятных воздействий и 2) восприятие пыльцы. Первая функция является общей для всех (не мужских) цветков с небольшими модификациями. Вторая функция характеризуется большим разнообразием. Она очень слабо развита у самоопыляющихся растений. При абиотическом опылении защитная функция околоцветника может стать препятствием для переноса пыльцы; следовательно, с точки зрения экологии опыления околоцветник у таких растений - негативный фактор. И, наконец, у растений, опыляемых животными, вторая функция цветка включает привлечение опылителей - обычно с помощью околоцветника. Рассмотрим эти три случая отдельно.
При рассмотрении процесса опыления необходимо ввести понятие единицы опыления (см. Berg, 1959), для которой мы предлагаем термин "anthium" У таких растений, как Tulipa или Paeonia, единицей опыления служит отдельный цветок: он привлекает насекомых, даже если он совершенно один. Опылители опускаются на цветок, т. е. они перестают двигаться и ограничивают свои движения только теми, которые необходимы для добывания аттрактантов: пыльцы, нектара и т. д. Совершенно ясно, что единицей опыления у Chrysanthemum leucanthemum, Trifolium pratense, Corrtus или Zanthedeschia является целое соцветие. Опустившееся на него насекомое перестает перемещаться (и использовать для этой цели энергию), и до тех пор, пока оно не оставит соцветие, все его движения в сущности не отличаются от вторичных движений, производимых насекомым, использующим отдельные нектарники индивидуального цветка. В экологии опыления морфологические различия между одиночными цветками и соцветиями не имеют значения; единицей опыления может быть как цветок, так и соцветие. Однако встречается много промежуточных типов, менее ясных, чем те, о которых только что упоминалось. В частности, на большом расстоянии соцветие может показаться одной единицей опыления, а при ближайшем рассмотрении оказывается, что оно представляет несколько единиц. Соцветие Linaria vulgaris можно определить как элемент аттрактации, но в этом случае движения опылителя, добывающего нектар из отдельных цветков, настолько отличны от движений опылителя, производимых при перемещении от одного нектарника к другому, что их следует рассматривать как перемещение, даже если иногда насекомое ползает, а не летает. Часто можно наблюдать, как в таких случаях насекомое перелетает с одного соцветия на другое, часто возвращаясь к тому, на котором оно уже побывало. Между прочим, такие бессистемные движения опылителей увеличивают возможности перекрестного опыления. Цветок Iris состоит из одного элемента аттрактации, но трех отдельных единиц опыления, также четко разделенных, как цветки Linaria, а у Gloriosa rothschildiana один цветок образует даже шесть единиц опыления (пять - мужских и одну - гермафродитную), рыхло соединенных в один элемент аттрактации. Иногда элемент аттрактации включает органы, которые не входят в единицу опыления, как, например, парус в околоцветнике Iris, прицветники Castilleja или цветоножки Mesadenia. Очевидно, что эти органы привлечения могут происходить как из той части цветка, для которой привлечение является "нормальной" функцией, т. е. из околоцветника, так и из других органов, выполняющих эту функцию частично или полностью, как, например, в двух последних родах, отмеченных выше. Следует при этом отметить, что если венчик Castilleja почти незаметен со стороны, то соцветия Mesadenia имеют относительно яркие прицветники, венчики и цветоножки.
В дальнейшем тексте мы будем употреблять "flower" как морфологический термин, a "blossom" (Percival, 1955) для элемента аттрактации как экологический термин. Это соответствует разнице между немецкими словами Blüte и Blume (см. Knoll, 1926), за исключением того, что мы включаем в "blossom" также отдельные цветки Iris и т. д., не упомянутые Кноллем, и не согласны с исключением элементов аттрактации для анемофильных растений (Knoll, 1956). Таким образом, "blossom" может быть либо соцветием (pseudanthium, Überblüte), цветком (euanthium, Blüte) или частью цветка (meranthium, Teilblüte, см. рис. 2).
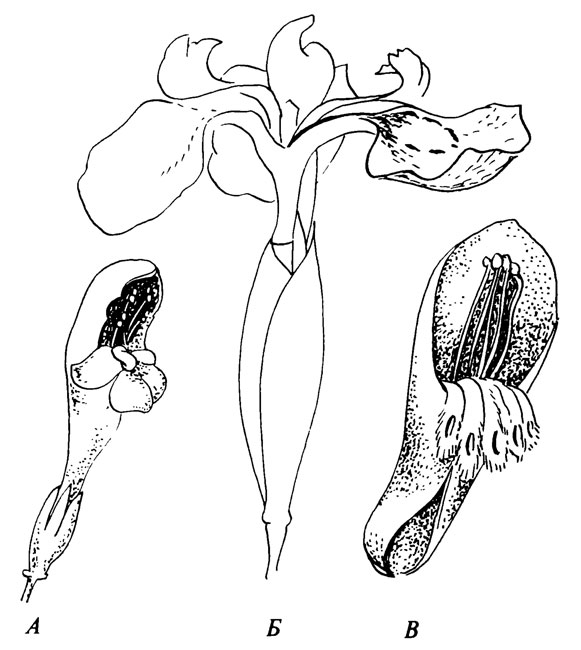
Рис. 2. Элемент аттрактации, образованный по типу цветка с зевом тремя разными способами. А. Весь цветок (Galeopsis). Б. Часть цветка, образуемая рыльцем и одним лепестком околоцветника (Iris pseudacorus). В. В соцветии этот элемент образован брактеей (верхней) и частями околоцветника отдельного цветка (Mimetes hartogii)
Простейшим элементом аттрактации можно считать открытый недифференцированный цветок, который предоставляет опылителю пыльцу (и нектар?) и рыльце пассивно, обычно это верхушечный цветок на главной или боковой оси. У ветроопыляемых растений развитие идет в направлении редукции околоцветника и увеличения количества пыльцы и размеров рыльца одновременно с более эффективным представлением обоих. У некоторых видов развивается активное выбрасывание пыльцы.
У растений, опыляемых животными, направление развития приводит к более высокой степени интеграции и точности в предоставлении, транспорте и распределении пыльцы.
Мы можем предполагать, что микроспоры первоначально возникли на примитивных, неспециализированных структурах и только позднее появились типичные тычинки, которые сначала имелись в большом количестве. В процессе эволюции число тычинок уменьшилось, и их местоположение определилось точнее. Эта линия развития достигла высшей точки в образовании поллиниев с пыльцевыми зернами, которые у Asclepiadaceae сливаются до такой степени, что имеют одну общую пору прорастания (ср. Jørgensen, 1919; Linskens, Suren, 1969).
Подобная линия развития может быть прослежена и для функции отложения пыльцы: а) свободные, открытые плодолистики с маргинальным рыльцем, столбик отсутствует (Drimys, Nypa, Akebia); б) свободные закрытые плодолистики со столбиком и рыльцем, в) плодолистики временно соединяются с рыльцем (Rutaceae, Ficus); г) слияние до образования синкарпных гинецеев. Такая интеграция может стать явной в процессе развития плода, когда апокарпные гинецеи дают синкарпные плоды, например у многих магнолиевых. При этом наблюдается тенденция от диффузного центрального расположения к точно определенному, часто асимметричному расположению нескольких плодолистиков.
Если обе функции присущи одному цветку, то конечным результатом эволюции может быть полное слияние как у Оrchidaceae, Asclepiadaceae или у Pedilanthus, причем нектарники и пыльники располагаются в глубоких узких частях цветка; такой же эффект может быть достигнут за счет того, что цветок открывается только тогда, когда насекомое коснется его. Примером этого последнего случая могут служить эфемерные растения. Дальнейшая эволюция шла по линии развития ловушек, закрытых цветков, зигоморфности, различий между частями цветка, введения проводниковых структур либо механических, подобных волоскам и колючкам, либо оптических, либо обонятельных, как, например, указатели нектара. Другим усовершенствованием явилось развитие активно движущихся частей цветка, подобных рыльцу Mimulus, тычинкам Centaurea, Cytisus или Catasetum.
В заключение следует сказать, что в обеих группах растений (опыляемых ветром и животными) наблюдается тенденция к объединению цветков в соцветия. Кроме того, можно говорить о тенденции к переходу от аллофилии (когда опыление может быть произведено любым опылителем) к эуфилии (когда опыление производится только очень специализированными, иногда уникальными, агентами). В соответствии с этим направлением в развитии растений шел процесс эволюции и у опылителей - от дис- и аллотропных к эутропным организмам, причем это можно проследить как между крупными таксономическими единицами - от жуков до чешуекрылых, так и внутри каждой группы. Подобное же развитие наблюдалось и в таксономических группах позвоночных (ср. табл. 2).
В экологии опыления существенное значение придается движениям растений под действием ветра (и воды для водных растений); под действием опылителя растения также могут производить более или менее случайные движения. Однако есть целый ряд очень специфических движений целого цветка, осуществляемых им для контакта с опылителем. Спонтанные и индуцированные движения помимо закрывания их под действием неблагоприятных условий могут изменить общую структуру цветков. Некоторые цветки в процессе цветения почти полностью меняются, захватывая своих опылителей с помощью различных структур, особенно с помощью ловушек (Calycanthus, Nymphaea, Victoria). Отдельные части некоторых цветков после того, как на них опустится насекомое, двигаются таким образом, чтобы создать условия, гарантирующие опыление. Нижняя губа у цветков многих орхидных способна производить ряд движений: очень простые, пассивные движения под действием тяжести опылителя (Epipactis palustris, Bulbophyllum); более сложные (Plocoglottis) и очень сложные, активные (сейсмонастические) движения, вызываемые касанием насекомого какой-то чувствительной части цветка (Pterostylis). В семействах Labiatae и Papilionaceae встречаются взрывающиеся цветки.
Каждая отдельная часть любой единицы опыления имеет свою функцию, но функции всех частей внутри этой единицы тесно связаны между собой. Опыт показывает, что определенные комбинации и определенные структуры встречаются чаще, чем другие, что приводит к образованию определенных типов единиц опыления. Для каждого из этих типов характерен также определенный механизм опыления. Однако это вовсе не означает, что все "типичные" черты присущи всем типам; существует определенный синдром, включающий признаки цветка, соответствующие определенному типу опыления, но в любом конкретном случае тот или иной признак, относящийся к синдрому, может отсутствовать в рассматриваемой единице опыления. Так, если установлено, что цветки, опыляемые летающими насекомыми (мухами), имеют тусклую неяркую окраску, это только означает, что статистически большая часть цветков, опыляемых мухами, имеет такую окраску, однако не исключено, что в отдельных случаях мухи опыляют яркоокрашенные цветки. Сходным образом, такой признак, как длинный узкий клюв, характерный для синдрома цветок - птица, замещается в случае опыления попугаями на короткий толстый изогнутый клюв с чрезвычайно специализированным языком. У гемитропных цветков синдромы бывают слабо развиты, но утрата каких-либо свойств может способствовать развитию новых адаптаций и видообразованию. Вместе с тем наличие всех признаков синдрома вовсе не является обязательным условием для успешного опыления; в конце концов в экспериментальных условиях колибри может получать пищу из рожка, а бесчисленное количество пчел воспользуется пищей, помещаемой на цветных квадратах и треугольниках. Однако постоянная встречаемость в природе таких комплексов признаков, составляющих синдром, свидетельствует о том, что это не является чем-то случайным или излишним. С другой стороны, поспешному исследователю не следует довольствоваться определением одного-единственного признака синдрома. Такой подход к проблеме синдрома из-за преждевременных заключений, основанных на слишком слабом доказательстве, бессмыслен.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'