Глава 3. Рассеивание спор и опыление
Опыление присуще только высшим растениям (т. е. растениям, обладающим пыльцой), однако на самом деле специализированный процесс такого типа характерен для большей части растительного царства. Если не принимать во внимание это положение, то процесс опыления представляется более сложным, чем это есть на самом деле. Для того чтобы правильно понять процесс опыления, его нужно рассматривать в связи с соответствующими явлениями, у низших растений. Как известно, фанерогамное растение представляет собой диплоидное поколение - спорофит. Само название свидетельствует о том, что это поколение образует споры, причем в процессе образования спор обычно происходит редукция хромосом. Споры дают гаплоидное поколение - гаметофиты (мужской или женский), образующие половые клетки, которые сливаются при оплодотворении; таким образом, вновь восстанавливается диплоидное число хромосом и образуется новое спорофитное поколение.
Схематически это можно представить на примере зеленой водоросли Ulva так:

Спорофит. Зеленая водоросль Ulva
У Ulva все споры имеют одинаковые половые потенции; для других растений, например для Selaginella, характерны два типа спор, образующих гаметофиты двух различных типов:
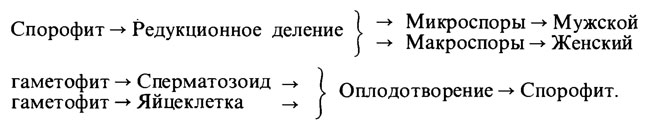
Для Selaginella, характерны два типа спор, образующих гаметофиты двух различных типов
Существование спор двух типов требует двух способов их распространения. Опыление связано только с распространением микроспор.
У цветковых растений также образуются макроспоры и микроспоры, но макроспоры находятся в пестике и никогда не отделяются от растения. Такое расположение макроспор делает микроспоры (пыльцевые зерна) более значительными, чем они есть на самом деле, вследствие кажущегося отсутствия соответствующего женского "двойника". Фактически молодое пыльцевое зерно и типичная микроспора мало отличаются друг от друга, если не считать нескольких клеток гаметофита (большей частью абортивных). В широком биологическом смысле опыление представляет собой просто один из особых случаев распространения микроспор, а экология опыления - один из аспектов экологии распространения спор. Так как деления ядер происходят в пыльцевом зерне позднее, они не влияют на способ распространения и, следовательно, в данном контексте их можно не рассматривать.
Растения (за исключением самых примитивных форм) имеют две неподвижные стадии - спорофит и гаметофит, и две подвижные - споры и гаметы. Исходно между (зоо-)спорами и гаметами не было заметных различий, но постепенно в ответ на изменения условий среды изменялись и споры. Гаметы в процессе филогенетического развития не претерпевали значительных изменений, не считая того, что они сильно редуцировались и почти утратили самостоятельность. Однообразие гамет приводит к тому, что процесс оплодотворения также довольно однообразен во всем растительном царстве. Даже у покрытосеменных процесс оплодотворения в своих главных чертах незначительно отличается от более примитивного процесса оплодотворения, характерного для низших растений.
Если гаметы ограничены в своем существовании первоначальной водной (или по крайней мере влажной) средой, в которой обитают примитивные растения, то споры приспособлены к распространению в сухих условиях. Жизнь растений на суше предполагает наличие спор, приспособленных к перенесению неблагоприятных воздействий окружающей среды. Именно поэтому споры даже наиболее примитивных наземных растений имеют прочную наружную оболочку, близкую по химическому составу к оболочке пыльцевых зерен. Таким образом, в процессе эволюции высшие растения использовали для своего опыления существовавший уже тип спор и уже развившийся механизм их распространения.
Однако в одном отношении между распространением спор и опылением наблюдается очень большое различие. Обычная спора имеет относительно широкую экологическую нишу; даже если гаметофит предъявляет строгие требования к среде, спора может прорасти во многих местообитаниях. К тому же, будучи подвижными, гаметы могут в некоторой степени компенсировать неблагоприятное взаимное положение женского и мужского половых аппаратов. Пыльцевое же зерно может прорасти, а развившийся из него гаметофит может расти лишь в одном - единственном месте, а именно на поверхности рыльца совместимого цветка. Если же прорастание происходит в каком-либо необычном месте, то пыльцевая трубка не только не может выполнить свою биологическую функцию, но просто не сможет далее развиваться.
Это сильно ограничивает способность пыльцевых зерен к прорастанию и требует исключительной точности в распространении по сравнению с точностью распространения простых спор. Многие замечательные адаптации (см. разд. 5.1), наблюдаемые специалистами в области экологии опыления, можно понять только исходя из этого требования точности. Эволюция механизмов опыления энтомофильных цветков представляет собой в основном эволюцию, направленную на достижение все более высокой точности.
В процессе эволюции гетероспории у многих низших растений также устанавливаются определенные требования к точности переноса микроспоры, причем в большей или меньшей степени изменяется подвижность "мужских" клеток.
У семенных растений одну из главных экологических функций споры низших растений, а именно распространение, принимает на себя совершенно новое образование - семя, представляющее собой стадию остановившегося в развитии нового спорофита. Семена имеют более широкую экологическую область распространения, чем пыльца, поэтому механизмы распространения семян обычно менее развиты (хотя не всегда менее разнообразны) и менее точны, чем механизмы опыления.
Для правильного понимания и оценки механизмов опыления перечислим сначала основные черты, характерные для рассеивания спор у низших растений. Проследить эволюционное развитие механизмов опыления на ископаемых растениях чрезвычайно трудно. Тем не менее ископаемые остатки дают нам кое-какие сведения; так, Крепет (Crepet, 1979а) пришел к выводу, что уже в юрский период наблюдалось взаимодействие между насекомыми и стробилами Cycadeoidea; эти стробилы несли и макро- и микроспорангии и обладали существенными преимуществами в синдроме опыления насекомыми. В других случаях соцветия, подобные соцветиям Fagaceae, в большей степени свидетельствуют об анемофилии. Крепет (Crepet, 1979 а, 1979b) заключил, что кантарофилия, миофилия, мелиттофилия и психофилия существовали уже в среднем эоцене.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'