Глава 3. Влияние температуры и водного дефицита на растения (засухоустойчивость растений) (Л. Г. Косулина)

Засухоустойчивость - способность растений в процессе онтогенеза переносить перегревы и обезвоживание и осуществлять в этих условиях свой рост, развитие и воспроизведение благодаря наличию ряда свойств, возникающих в процессе филогенеза под влиянием условий существования и естественного отбора.
Проблеме устойчивости растений к засухе большое внимание уделялось в работах А. А. Измаильского (1863), В. В. Докучаева (1892) и К. А. Тимирязева (1893). Позже крупный вклад в эту проблему внесли работы Н. А. Максимова с учениками и последователями. В последние годы вопросы засухоустойчивости широко обсуждаются в работах А. М. Алексеева, В. Ф. Альтергота, П. А. Генкеля, Н. А. Гусева, В. Н. Жолкевича, М. Д. Кушниренко, Н. С. Петинова, Н. А. Сатаровой, Дж. Левитта, Виэйра да Сильва и др.
Засуха является результатом действия высоких температур, а также водного дефицита. Для растений, произрастающих в северо-западных районах страны, где температура воздуха редко достигает +30°С, основной причиной засухи является дефицит влаги в среде их обитания (засуха северного типа). У растений, обитающих в южных и юго- восточных районах, нарушение водообмена проявляется в основном при действии высоких температур (засуха южного типа).
П. А. Генкель определяет засуху как биометеорологическое явление, которое характеризуется длительным или кратковременным бездождевым периодом, повышенной температурой воздуха, увеличением дефицита насыщения влажности воздуха, что вызывает усиление транспирации, в результате чего происходит обезвоживание и перегрев растений, которые могут привести к повреждению, снижению продуктивности и, возможно, гибели растений [5].
Различают засуху атмосферную и почвенную. Атмосферная засуха возникает при недостаточной увлажненности и высокой температуре воздуха. При атмосферной засухе у растений усиливается транспирация, что может привести к чрезмерной потере ими воды. Если температурные условия среды находятся в пределах толерантной зоны и корневая система растений хорошо развита, то непродолжительная атмосферная засуха не причиняет растительным тканям большого вреда. Продолжительная атмосферная засуха приводит к почвенной засухе, которая более опасна для растений.
Почвенная засуха в основном наблюдается в середине или конце лета, когда зимние запасы влаги исчерпаны, а летних осадков недостаточно. Происходит нарушение водного режима растений, что в разной степени отражается на всех физиологических функциях. Засуха способствует выработке целого ряда приспособительных признаков. Баланс между добыванием и расходованием воды достигается путем регулирования процессов поглощения и испарения воды. Засухоустойчивость у одних растений обусловлена приспособлениями к недостатку воды в атмосфере, у других - в почве.
В природе комбинация дефицита влаги в почве, атмосфере и высоких температур по-разному влияет па растения. Засуха северо-западного типа характеризуется низкой влажностью воздуха и почвы при умеренной температуре. Засуха юго-восточного типа характеризуется, как правило, пониженной влажностью воздуха и почвы в сочетании с высокими температурами. Следует отметить засуху мерзлотную, которая определяется действием низких температур и низкими значениями влажности почвы и воздуха.
По терминологии Г. Вальтера, весь растительный мир по отношению к воде делится на две группы: пойкилогидрические и гомойогидрические растения.
Пойкилогидрические растения не способны регулировать свой водообмен. К этой группе относятся многие энифитные и почвенные водоросли, лишайники, некоторые мхи, папоротники, папоротникообразные и некоторые покрытосеменные растения. Среди них можно выделить три экологические подгруппы: пойкилогигрофиты, нойкиломезофиты и нойкилоксерофиты. Все они не регулируют свой водообмен, но гигрофиты и мезофиты при сильном обезвоживании погибают, а ксерофиты впадают в анабиоз (криптобиоз). В этом состоянии у пойкилоксерофитных форм прекращаются все обменные процессы, а сохраняется лишь потенциальная способность к обмену, и при наличии благоприятных условий происходит возобновление обменных процессов.
Установлено, что только те растения, которые способны сохранять энергетически полноценное дыхание до самого конца обезвоживания, могут гелефицировать свою цитоплазму и в течение длительного времени в условиях засухи сохранять жизнеспособность. Зерновки пшеницы сохраняют ее в течение 20 - 25 лет, а семена лотоса, находившиеся в иловых отложениях в анаэробных условиях, сохраняют всхожесть на 75% в течение почти 2000 лет. Эти семена имеют гелефицированную цитоплазму и находятся в состоянии анабиоза.
Обезвоживание для пойкилоксерофитов является не патологией, а нормой реагирования. Основным признаком этой группы растений является способность сохранять энергетическую полноценность дыхания. У этих растений кислоторастворимый органический фосфор, содержащий макроэрги, не разрушается при обезвоживании. Пойкилоксерофитные формы сохраняют способность к синтезу белка в процессе обезвоживания. Даже после 10-месячного пребывания мха Necker в анабиозе полирибосомы сохраняют способность к синтезу белка, и через 10 - 30 мин. после перенесения их во влажную среду у них обнаруживается этот процесс.
Реакция основных физиологических процессов на обезвоживание у пойкилоксерофитных растений в основном протекает однофазно - интенсивность дыхания и фотосинтеза постепенно снижается по мере потери ими воды. Кроме того, при водном дефиците у них не происходит существенных изменений в гормональном балансе, в частности, не накапливаются ингибиторы, что позволяет данным организмам быстро восстанавливать ростовую функцию сразу же после их увлажнения.
Гомойогидрические растения - это покрытосеменные высшие растения, которые регулируют свой водный режим. Некоторые из них способны выносить значительное обезвоживание, но оно не доходит до воздушно-сухого состояния, и эти растения не способны впадать в анабиоз. Этой способностью обладают только семена.
Соотношение между добыванием и расходованием воды у гомойогидрических растений складывается различным образом. Это зависит от многих факторов, в том числе от способности растений приспосабливаться к различным экологическим условиям. По этому признаку гомойогидрические растения делят на гигрофиты, мезофиты и ксерофиты.
Наиболее засухоустойчивыми являются растения засушливых мест обитания - ксерофиты (от греч. xerox - сухой). Характерным признаком их является наличие различных приспособлений для сохранения воды в клетках. Группа ксерофитов неоднородна, и по характеру адаптации к засухе выделяют следующие типы (по П. Л. Генкелю).
1. Суккуленты (по Н. А. Максимову - ложные ксерофиты) - растения с толстыми мясистыми тканями, имеющими большой запас воды (кактусы, алоэ, агава, молодило и др.). Вода концентрируется у них в листьях или стеблях, покрытых толстой кутикулой, волосками. Они имеют небольшое количество устьиц, которые большую часть дня закрыты. Для суккулентов характерен САМ - тип фотосинтеза. Они имеют слабо развитую, поверхностную корневую систему, которая максимально использует выпадающие осадки, а также высокую водоудерживающую способность биоколлоидов протоплазмы. Эти растения отличаются очень экономным расходованием воды. Суккуленты плохо переносят длительное обезвоживание. В период сильной засухи у кактусов отмирают боковые и остается центральный корень. Эти растения характеризуются медленным ростом.
Суккулентность в основном связана с разрастанием клеток паренхимы при одновременном увеличении вакуоли и сильном сокращении размеров межклетников. Существуют листовые (агава, алоэ и др.), стеблевые (кактусы) и корневые (сейба мелколистная) суккуленты. На корнях сейбы мелколистной образуются вздутия диаметром до 30 см, и в период дождей в их губчатой сердцевине накапливается влага. Эти "корневые хранилища влаги" могут функционировать много лет.
2. Настоящие ксерофиты (эвксерофиты). К ним относятся такие степные растения, как вероника сизая, полынь голубая, верблюжья колючка и др. Эвксерофиты имеют небольшие тонкие листья с большим количеством устьиц, хорошо развитую корневую систему. Высокий осмотический потенциал их клеток позволяет этой группе растений поглощать не только легко-, но и труднодоступную воду. Для них характерна хорошо развитая проводящая система и интенсивная транспирация.
3. Стипаксерофиты - степные злаки (ковыль, типчак, перекати-поле и др.) Они имеют жесткие кожистые листья с большим числом устьиц и слаборазвитую корневую систему. В период засухи листья этих растений скручиваются, и устьица оказываются внутри трубочки. Кроме того, клетки данных растений имеют высокий осмотический потенциал, что позволяет им добывать труднодоступную воду. Они быстро используют влагу летних дождей. Эта группа растений способна переносить длительные перегревы.
4. Полуксерофиты (гемиксерофиты - шалфей, резак обыкновенный, арбуз, груша и др.) не способны переносить длительное обезвоживание и перегрев. Они обладают интенсивной транспирацией и низкой вязкостью протоплазмы. Гемиксерофиты имеют глубокую корневую систему, которая может достигать подпочвенных вод, что и обеспечивает бесперебойное снабжение их влагой.
Существуют растения, у которых дефицит воды в значительной степени определяется холодными почвами.
1. Психрофиты (сосна сибирская, ель, береза, черника, багульник и др.) произрастают на увлажненных и холодных почвах в зоне тундры. Из-за низких температур почвы вода для данных растений физиологически недоступна. У них хорошо развита ксероморфная структура. Для игл хвойных древесных пород характерны погруженные устьица, которые закрыты зимой смоляными пробками, а также толстостенный эпидермис с подстилающей его гиподермой. Все это служит для уменьшения потерь воды у исихрофитов в летнее и зимнее время.
2. Криофиты (подушки, произрастающие в Забайкалье) - растения сухих и холодных местообитаний - характеризуются замедленными темпами водообмена и низкой оводненностью тканей.
П. А. Генкель выделил среди ксерофитов подгруппы: лауроксерофиты - вечнозеленые растения Средиземноморья; аэроксерофиты - растения, которые растут за счет атмосферной влаги, в частности туманов [5].
Мезофитные растения (от греч. mesos - средний, промежуточный) произрастают в умеренном климате средних широт, менее устойчивы к засухе по сравнению с ксерофитами. К ним относятся хлебные злаки и ряд сельскохозяйственных растений (горох, огурцы, табак, салат и др.), древесные растения, кроме ольхи и некоторых видов ивы. Их устойчивость к засухе обусловлена тем, что эти растения способны регулировать интенсивность транспирации за счет работы устьичного аппарата, сбрасывания листьев и др. Для них характерна хорошо развитая корневая система, развивающая достаточно высокое корневое давление. Высокий уровень осмотически активных веществ в клетках (углеводов, органических кислот, минеральных ионов) обусловливает значительную водоудерживающую способность растительных тканей.
Мезофитные растения более требовательны к воде, чем ксерофиты. У мезофитных растений чем выше по стеблю расположен лист, тем мельче его клетки, гуще сеть проводящих пучков, сильнее развита палисадная паренхима. Устьица у листьев верхних ярусов даже при водном дефиците дольше остаются открытыми. Это способствует более длительному поддержанию у них процессов фотосинтеза и увеличению концентрации клеточного сока, что позволяет им оттягивать воду от ниже расположенных листьев. Мезофитные растения очень разнообразны по своим физиологическим и анатомическим признакам. Многие из мезофитов обладают значительной пластичностью метаболических процессов, что позволяет им изменяться в направлении ксерогенеза, связанного не с потерей продуктивности, а с ее повышением в аридных условиях. Ксеромезофиты обитают в районах с периодически возникающим водным дефицитом. Адаптация этих растений связана с развитием глубокопроникающей корневой системы. Благодаря этому различные виды дуба встречаются в степях и даже в полупустынях. Из травянистых растений к ксеромезофитам относятся отдельные виды клевера, люцерны, мятлика и др.
Кроме того, существуют мезофитные растения, которые произрастают на почвах с избыточным увлажнением и имеют поверхностную корневую систему. Это гигромезофиты (недотрога, звездчатка и др.), у которых почти не развита система межклетников.
Гигрофитные растения (от греч. nygros - влажный) обитают в районах избыточного увлажнения с большим количеством осадков и высокой влажностью воздуха. К ним относятся растения, развивающиеся под пологом густого леса (кислица и другие тенелюбивые травы), растения открытых сырых почв - калужница, чистяк, сердечник и многие осоки. У них нет приспособлений, ограничивающих расход воды. Они имеют крупные листовые пластинки с тонкой кутикулой и немногочисленными устьицами. У них почти отсутствует устьичная транспирация, и они используют кутикулярную. Клетки у данных растений достигают значительных размеров, имеют тонкостенные оболочки. Для гигрофитов характерны слабо одревесневшие стенки сосудов, плохо развитые механические ткани. Эти растения имеют, как правило, длинные стебли и недостаточно развитую корневую систему. Осмотический потенциал их клеток невысок. Незначительный недостаток воды в почве вызывает быстрое завядание гигрофитов. У них хорошо развиты приспособительные реакции к гипоксии.
По условиям произрастания и особенностям строения к гигрофитам очень близки растения с частично или полностью погруженными в воду или плавающими на ее поверхности листьями, которые называются гидрофитами.
В связи со спецификой условий существования основной структурной особенностью гидрофитов является губчатость, "рыхлость" их тканей, наличие крупных межклетников, полостей, заполненных воздухом, т. е. преобладание аэренхимы. Полости погруженных в воду частей растений обычно сообщаются с устьицами, расположенными на непогруженных листьях, что способствует процессу газообмена и проникновению кислорода к органам, расположенным в среде с небольшим содержанием кислорода. Губчатость тканей и воздушные полости в листе и черешке поддерживают их плавучесть. Для гидрофитов характерна атрофированность поддерживающих, механических тканей. Отсутствие кутина и суберина у погруженных органов способствует поглощению воды и питательных веществ непосредственно поверхностью органа - дополнительно к тому, что растения могут получать через корни.
У погруженных водных растений (валлиснерии) отсутствует транспирация. Поддержание восходящего водного тока у прикрепленных (укореняющихся в донном грунте) гидрофитов осуществляется благодаря корневому давлению и деятельности гидатод - специальных образований, через которые происходит выделение воды листьями (процесс гуттации).
У гидрофитов с плавающими на поверхности воды листьями (у кувшинок) наблюдается интенсивная транспирация. Они не выносят даже небольшого обезвоживания. Недостаток механических тканей компенсируется у них плавучестью отдельных органов, в частности листьев.
Гелофиты (стрелолист, тростник и др.) - растения береговых и прибрежных местообитаний с избыточным увлажнением. Они способны переносить как значительное обезвоживание, так и временное затопление.
Продолжительная засуха вызывает у некоторых мезофитных и ксерофитных растений различного вида нарушения в обмене веществ: 1) ухудшается водный режим с появлением дефицита влаги, что вызывает торможение ростовых процессов, нарушение формообразования, повреждение фотосинтетического аппарата и снижение фотосинтеза; 2) увеличивается интенсивность дыхания, но оно оказывается непродуктивным, т. е. происходит разобщение окисления и фосфорилирования; 3) снижается вязкость протоплазмы, ее эластичность; 4) снижается синтетическая деятельность корней (уменьшается содержание органического фосфора и азота); 5) повреждаются мембранные компоненты протоплазмы и повышается проницаемость; 6) снижается содержание суммарной РНК, общего и белкового азота; 7) соотношение между синтезом и гидролизом сдвигается в сторону гидролиза, увеличивается активность гидролитических ферментов (в частности, рибонуклеазы); 8) повреждаются наиболее чувствительные к засухе генеративные органы, что ведет к неполному плодоношению. В результате в условиях длительной и повторяющейся засухи снижается общая продуктивность.
Засухоустойчивость разных органов растений неодинакова. Молодые растущие листья за счет притока ассимилятов относительно более устойчивы, чем листья, закончившие свой рост. В условиях длительной засухи отток веществ в молодые листья может происходить и из генеративных органов. Засухоустойчивость зависит также и от фазы развития растений. Яровые и озимые злаки (пшеница, рожь) наиболее чувствительны к недостатку влаги в фазы выхода в трубку и колошения, кукуруза - в период цветения и молочной спелости, просо и сорго - в фазы выметывания метелки и налива зерна, зернобобовые - в период цветения, картофель - цветения и формирования клубней, подсолнечник - образования корзинки и цветения, хлопок - в фазу цветения и заложения коробочек, бахчевые в период цветения и созревания.
У дикорастущих растений в борьбе с засухой выработался целый арсенал средств защиты, многие из которых присущи и культурным растениям: устьичная регуляция, строение корневой системы и листьев. Однако у культурных растений эти свойства менее выражены. Создавая сорта, селекционеры стремились вывести высокоурожайные формы, не учитывая, что урожай - это компромисс между продуктивностью и устойчивостью в неблагоприятных условиях. Поскольку этот компромисс (урожай и устойчивость) пока не удается устранить, селекционеры стремятся использовать его с максимальной выгодой, жертвуя той частью растений, которая менее ценна. С этой целью создавались короткостебельные сорта пшеницы, которые отличаются высокой продуктивностью, но они незасухоустойчивы - требуют орошения, хорошо отзываются на удобрения.
Значительная стойкость растений к засухе определяется за счет 1) адаптивных морфологических особенностей стебля, листьев, генеративных органов, повышающих их выносливость, толерантность; 2) ксероморфной структуры и хорошо развитой корневой системы; 3) повышения вязкости, эластичности и водоудерживающей способности протоплазмы и содержания связанной воды в клетках; 4) функциональной сохранности клеточных структур и синтетических процессов на высоком уровне; 5) приспособления роста и развития к условиям среды.
К. А. Тимирязев в своих исследованиях уделял большое внимание приспособительным признакам у растений. Среди них он отмечал различные формы волосков на листьях, создающих опушенность листовой поверхности и точно войлок защищающих живые клетки, заглубление устьиц и их расположение на нижней стороне листа, ориентацию самих листьев, свертывание их при высокой температуре и при недостатке влаги. Глубинные устьица могут быть открыты в самые жаркие часы дня, что важно для фотосинтеза, и в то же время не происходит повреждения растений в этих условиях [30].
Большой вклад в проблему засухоустойчивости внес В. Р. Заленский, который обнаружил существенные различия по строению и по отношению к засухе между листьями разных ярусов у травянистых и древесных растений. Отмечается ксероморфное строение верхних (молодых) листьев - меньшая величина клеток и межклетников, клетки эпидермиса снабжены плотной оболочкой и толстым восковым покровом. Верхние (молодые) листья имеют большое количество устьиц и обладают значительной транспирацией. Они, что очень существенно, имеют большую сосущую силу и, следовательно, легко отнимают воду от нижележащих листьев и таким образом удовлетворяют свои потребности в воде. Следовательно, большая засухоустойчивость верхних листьев сочетается с меньшим содержанием воды в них и с более интенсивной транспирацией. Эти положения (закон Заленского) долго оставались без внимания со стороны физиологов, так как господствовала точка зрения, что засухоустойчивость связана с более экономным расходованием воды [11].
В дальнейшем Н. А. Максимов подтвердил эти положения и показал, что, несмотря на защитные приспособления, транспирация у ксерофитов оказалась значительно выше, чем у мезофитов. Мезофиты расходуют чаще всего 3 - 4 мг воды в час на единицу (см2) листовой поверхности, а ксерофиты за то же время и с той же площади листа - 8 - 14 мг воды [20].
Б. А. Келлер показал, что у ксерофитов на солнце транспирация значительно интенсивнее, чем у мезофитов, даже в пределах одного рода. Если у ксерофита (Ясменник сизый) интенсивность транспирации принять за 100%, то у мезофита (Ясменник пахучий) она составляет 38% [14].
Исследования В. Р. Заленского, Б. А. Келлера, Н. А. Максимова показали также, что у ксерофитов на единицу площади листа приходится значительно больше устьиц и у них лучше развита проводящая система, чем у гигрофитов [3; 18].
Таким образом, засухоустойчивость следует рассматривать как способность клеток активно добывать воду и обеспечивать свои ткани водой в наиболее трудных условиях.
По мнению Дж. Левитта, засухоустойчивость растений обеспечивается за счет уклонения их от засухи, избегания обезвоживания и повышения степени выносливости к обезвоживанию [34].
Растения засушливых южных областей выработали специфические защитные свойства, позволяющие им снижать вредное влияние засухи и перегрева. Так, существует группа растений, которые уклоняются от засухи. Это - эфемерные растения (маки, вероники и др.), не обладающие ксероморфной структурой и развивающиеся только во время короткого периода дождей, когда они в течение нескольких недель достигают зрелости. После этого их активная жизнь прекращается до будущего года. Эфемеры - однолетние растения, сохраняющиеся в сухое время только в виде семян. Эфемероиды - многолетние растения, сохраняющиеся в сухое время в виде луковиц, клубней, корневищ (вероники, тюльпаны и др.).
Эфемеры и эфемероиды составляют до 70% видов пустыни Каракумы.
Некоторые ксерофиты избегают перегрева, интенсивно испаряя влагу, и имеют глубокозалегающую корневую систему с высокой сосущей силой. Ряд растений, обитающих в пустыне, обладают корнями, проникающими в почву на 20 - 30 м и разветвляющимися только вблизи грунтовых вод. Эти растения не имеют приспособлений для уменьшения испарения воды. В Сахаре, где подпочва скалиста, распространен другой тип ксерофитов. Могучая корневая система этих растений развивается в горизонтальном направлении, распространяется но громадным массивам почвы и поглощает после дождя за короткое время значительное количество воды.
У других ксерофитов во время засухи закрываются устьица, листья скручиваются. Ущерб, который испытывают эти растения, компенсируется высокой активностью фотосинтетического аппарата. Установлено, что ксерофиты фотосинтезируют интенсивнее, чем мезофиты. У некоторых растений листья приобретают суккулентный тип, у других они редуцированы, а функцию фотосинтеза выполняет стебель. Эти растения слабо транспирируют и фотосинтезируют, но отличаются высокой засухо- и жароустойчивостью.
Не менее важным для перенесения неблагоприятных условий является обильное образование органов плодоношения, которые, находясь в состоянии покоя, могут сохранять жизнь в течение неблагоприятных периодов, обеспечивая сохранение вида. В экстремальных условиях засухи некоторые ксерофиты теряют листья летом, поэтому имеют два периода покоя.
Различные типы адаптации к засухе обнаружены не только у ксерофитов, но и у плодовых растений. Одни (груша) имеют глубокозалегающую корневую систему, интенсивную транспирацию, повышают при засухе водный потенциал листьев. Другие (яблоня, слива) имеют поверхностную корневую систему, высокую водоудерживающую способность клеток и повышенный осмотический потенциал при засухе. У персика при засухе повышается водный потенциал, в том числе и осмотический, преобладает содержание белков в листьях над углеводами. Клетки персика обладают высокой водоудерживающей способностью.
Развитие и выживание растений в условиях засухи в значительной степени зависит от обеспеченности клеток водой [25; 23; 10]. Действие засухи в первую очередь приводит к уменьшению в растениях свободной воды. Различие свойств свободной и связанной (осмотически и коллоидносвязанной) воды определяет и разное их значение в жизни растений. Уровень свободной воды обусловливает интенсивность физиологических процессов, связанной - устойчивость растений к неблагоприятным условиям среды. Значительное снижение содержания свободной воды в условиях атмосферной или почвенной засухи приводит к нарушениям физиологических процессов и в конечном счете - к повреждению растений. Известно, что количество связанной воды у ксерофитов значительно выше, чем у мезофитов. П. А. Генкель и М. Д. Кушниренко считают, что одним из показателей повышенной засухоустойчивости у мезофитных сельскохозяйственных растений и плодовых культур является высокое содержание связанной воды [5; 17].
Неблагоприятное действие засухи на растение зависит от степени, глубины и продолжительности влияния водного дефицита. При умеренной засухе происходит усиление взаимодействия белков с водой, т. е. увеличение количества связанной воды. Результатом чего является повышение стабильности цитоплазмы и увеличение водоудерживающей способности клеток. А это позволяет растениям переносить умеренную засуху.
При сильной засухе начинают преобладать нарушения структурной организации и функциональной активности клеток. (Содержание воды в клетках уменьшается, что может указывать на ослабление взаимодействия белков с водой.
Не касаясь вопросов структуры воды, следует сказать о трех типах воды в растении, которые имеют определенное значение для переживания засухи. К. А. Тимирязев отмечал существование в клетке организационной и расхожей (легкообмениваемой) воды. Б. Б. Вартапетян и А. Л. Курсанов выделили метаболическую воду [16]. Образование метаболической воды является одним из процессов приспособления мезофитов к засухе. Так, у растений томатов во время засухи в течение часа образуется примерно 13% воды за счет дыхания, даже если оно энергетически неполноценно. В 1982 г. П. А. Генкель и Н. А. Шеламова ввели понятие "гомеостатическая вода". Это наименьшее количество воды, при котором растение способно поддерживать постоянство своей внутренней среды (гомеостаз) [6]. Определенный уровень гомеостатической воды характеризует устойчивость растений к обезвоживанию. У ксерофитных организмов уровень гомеостатической воды достигает 25 - 27%, у мезофитов - 45 - 60%, у гигрофитов - 65 - 70% от массы сырого вещества.
Сущность адаптации растений к засухе сводится к обеспеченности клеток и тканей водой, к поддержанию структурной и функциональной целостности растительного организма, а это обеспечивает рост и образование репродуктивных органов в экстремальных условиях водоснабжения.
Известно, что водный обмен высших растений включает три этапа: 1) поглощение воды из почвы системой мелких корешков и корневых волосков с последующим поступлением ее в мертвые клетки ксилемы; 2) передвижение воды по ксилеме, иногда на расстояния, измеряемые десятками метров; 3) отдача воды в процессе транспирации.
Растения аридных местообитаний (пустынь) обладают широко расходящимися или очень глубоко проникающими в почву корнями, что позволяет им поглощать грунтовую воду. В этих областях и в засушливые периоды вода поднимается в почве под действием капиллярных сил. При диаметре почвенных частиц 0,2 - 0,1 мм высота капиллярного подъема составляет 50 см, а при диаметре 0,05 - 0,002 мм - 200 см.
Сухой воздух имеет очень низкий водный потенциал. При влажности воздуха 80% водный потенциал достигает значений порядка - 30 МПа, при 50% влажности воздуха - 100 МПа. В почве водный потенциал достигает высоких значений. В зависимости от типа и активности почв значения водного потенциала колеблются в широких пределах: от - 05 МПа до - 0,5 МПа. Растения занимают промежуточное положение в интервале значений водного потенциала воздух-почва (в корневых клетках -0,6÷-0,8 МПа, в надземных органах -1,0÷-1,5 МПа). Вода всегда движется в сторону более низкого водного потенциала.
Так как давление водяного пара в воздухе, как правило, сравнительно невелико, неизбежны потери воды у растений в результате транспирации. Кутикулярный слой на листьях практически непроницаем для водяного пара. Однако у молодых листьев растений с тонкой кутикулой в условиях засухи, когда устьица закрыты, интенсивность кутикулярной транспирации может достигать 50% от общей транспирации. В связи с этим молодые листья особенно чувствительны к водному дефициту. И наоборот, виды с толстым слоем кутина в кутикуле (виды магнолий и хвойных) теряют очень мало воды через эпидермис листа после того, как устьица закрываются. Вода может испаряться с поверхности ветвей и стволов древесных растений через чечевички и окружающие их слои пробки (периферическая транспирация). Общее количество воды, испаряемой через чечевички, значительно меньше потерь воды через листья. Однако вследствие транспирации ветвей в зимнее время часто возникает водный дефицит у древесных растений, и они могут погибнуть в результате обезвоживания.
Основная масса воды испаряется через устьичные щели. Движение устьиц находится под влиянием как внешних (влажности воздуха, условий водоснабжения, света и температуры), так и внутренних факторов (парциального давления CO2 в системе межклетников, состояния гидратации растений, ионного и гормонального баланса).
При значительной атмосферной и почвенной засухе устьица закрываются и таким образом предотвращается избыточная потеря влаги в данных экстремальных условиях. При потере влаги после засухи устьица долгое время остаются закрытыми, их движение определяется изменением водного потенциала растений и почвы. Закрывание устьиц в значительной степени определяется уменьшением в вакуолях замыкающих клеток содержания осмотически активных веществ (K+, Cl-, малата). Это вызывает усиление выхода воды из замыкающих клеток, снижение тургорного давления и, как следствие, закрывание устьичной щели. Этот процесс регулируется CO2 и абсцизовой кислотой. Закрывание устьиц происходит в результате повышения концентрации CO2 в межклетниках листа ночью, когда в результате отсутствия фотосинтеза и продолжающегося дыхания уровень CO2 в тканях повышается. У суккулентов закрывание устьиц происходит днем, когда при декарбоксилировании малата высвобождается углекислота, которая накапливается в межклетниках перед дальнейшем ее использованием. При высоких концентрациях CO2 внутриклеточная среда в замыкающих клетках закисляется, что снижает активность ФЭН-карбоксилазы. В результате снижения уровня малата в замыкающих клетках происходит уменьшение осмотического давления и закрывание устьиц.
Абсцизовая кислота выполняет роль экзогенного антитранспиранта. Водный дефицит стимулирует образование абсцизовой кислоты, которое происходит при определенном пороговом значении водного потенциала (для большинства растений от - 0,8 до - 1,0 МПа). Содержание АВК в тканях в среднем увеличивается на порядок со скоростью 0,15 мкг/г сырой массы в час. Под влиянием высоких концентраций АВК происходит перенос ионов калия из замыкающих клеток в соседние, эпидермальные. В результате этого в замыкающих клетках падает осмотическое давление и устьичная щель закрывается. АВК также способствует запасанию гидратной воды в клетке, так как активирует синтез пролина, увеличивающего оводненность белков в условиях засухи. Кроме того, под влиянием высоких концентраций АВК происходит увеличение проницаемости корневых клеток для воды. Следовательно, АВК обеспечивает поглощение воды корнем и затрудняет отдачу воды листьями, что в целом приводит к улучшению водного баланса в условиях засухи.
Таким образом, способность устьичного аппарата предотвращать избыточную потерю влаги при засухе является одним из путей адаптации растений к водному стрессу.
Кроме морфоанатомических особенностей листа (кутикула, восковой налет, опушение и т. д.), корней (степень их развития и расположения в почве), структуры и поведения устьичного аппарата, важную роль в противодействии обезвоживающему влиянию засухи играет водоудерживающая способность клеток. П. А. Генкель придает большое значение водоотдаче в качестве одной из важных характеристик засухоустойчивости растений. Он показывает разный характер изменений водоудерживающей способности листьев различных по засухоустойчивости сортов пшеницы и ячменя при прогрессирующей засухе. Вначале водоудерживающая способность клеток увеличивается у всех сортов, причем у неустойчивых сильнее. При достижении коэффициента завядания (потеря воды листьями 8 - 10%) у неустойчивых сортов данный показатель значительно снижается, а у устойчивых продолжает повышаться за счет усиления взаимодействия белков и воды.
По данным Н. А. Гусева при умеренной засухе (остаточный водный дефицит 10 - 15%) водоудерживающая способность листьев яровой пшеницы увеличивалась, при усилении засухи (остаточный водный дефицит 20 - 25%) - резко снижалась за счет увеличения проницаемости мембран. В то же время содержание связанной (прежде всего гидратной) воды оставалось примерно таким же, как при умеренной засухе [8].
При сильном обезвоживании клеток наблюдается увеличение проницаемости мембран, что связано с нарушением их структуры. Уже на ранних этапах обезвоживания происходят конформационные изменения фосфолипидных цепочек мембраны, уменьшается плотность расположения молекул и нарушается аллостерическая регуляция работы ряда мембранно-связанных ферментов. При дальнейшем обезвоживании наблюдается усиление активности кислых и щелочных липаз и изменение липидного состава мембран, приводящие к более существенным нарушениям в структуре мембранно-связанных ферментов. Когда содержание воды становится меньше 20% от первоначального веса мембраны, ее структурные и функциональные изменения зачастую принимают необратимый характер [26].
Барьерная роль мембран в транспорте воды отмечается в ряде работ других ученых. Так, Л. Х. Гордон показал, что проницаемость мембран для воды (10-4см/с) в 100 тыс. раз меньше скорости диффузии молекул воды через водный слой толщиной 75 А° [7].
Одним из мощных модификаторов и стабилизаторов мембраны является кальций, который контролирует ее проницаемость и водоудерживающую способность. Полагают, что это обеспечивается усилением - ослаблением (появлением - удалением) мостиков между фосфолипидами и белками. Вытеснение кальция из плазмалеммы с помощью анестетиков (пииольфена, аминазина) приводит к усилению выхода воды и ионов калия.
По мере обезвоживания проницаемость мембраны настолько нарушается, что анестетики не оказывают на нее влияния.
Следует отметить, что у засухоустойчивых растений при водном дефиците в меньшей степени изменяется проницаемость мембран по сравнению с неустойчивыми видами.
При значительной засухе важную роль в регуляции водоудерживающей способности листьев играют хлоропласты. Показана способность хлоропластов к самостоятельному регулированию своего водообмена: при слабом и умеренном обезвоживании клеток хлоропласты набухают, а при более сильном - хотя и теряют воду, но значительно медленнее, чем клетка в целом. В частности, при подсушивании в течение часа общая оводненность листьев кукурузы уменьшается на 2 - 6% (в зависимости от яруса), а оводненность хлоропластов, наоборот, возрастает на 21 - 30%. Даже при далеко зашедшем обезвоживании, когда оводненность клеток падает на 75%, оводненность пластид уменьшается меньше чем наполовину. Такое поведение хлоропластов при обезвоживании клеток, по-видимому, следует рассматривать в качестве приспособительной реакции фотосинтетического аппарата в условиях засухи. Таким образом, при слабом и умеренном обезвоживании клеток набухшие хлоропласты являются запасными резервуарами воды в клетке, а в условиях сильного водного стресса они - основные вместилища внутриклеточной воды. Меньшая потеря воды хлоропластами позволяет сохранять высокий уровень фотосинтеза в условиях засухи.
Водный режим растения оказывает влияние на процессы фотосинтеза. Вода непосредственно участвует в нем как субстрат окисления и источник кислорода. Величина оводненности листьев определяет степень открывания устьиц и, следовательно, поступление CO2 в лист. В условиях засухи, когда клетка теряет очень много воды, происходит закрывание устьиц вследствие изменения гормонального и ионного баланса. В 1949 г. В. А. Бриллиант обнаружила, что наибольшя фотосинтетическая активность у листьев наземных растений достигается обычно не при максимальном тургоре, а при небольшом дефиците воды, составляющем 5 - 6% от полного влагонасыщения клеток [4]. Как показывают исследования различных авторов, максимальный фотосинтез в листьях большинства наземных растений достигается при небольшом водном дефиците (~10%). Дальнейшее уменьшение оводненности листа приводит к значительному снижению фотосинтеза (рис. 10).
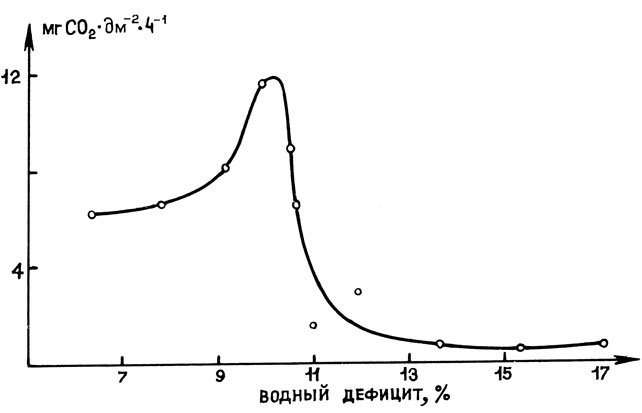
Рис. 10. Зависимость интенсивности фотосинтеза от водного дефицита ассимиляционной ткани у картофеля (по В. М. Иванченко, 1974)
В процессе эволюции появились разнообразные биотипы растений с фотосинтетическим аппаратом, приспособленным для эффективной работы в условиях засухи. Так, в районах с высокими температурами преимущество имеют C4-растения с высокой эффективностью фотосинтеза (ползучие сорняки). Максимальная скорость включения CO2 у этих растений может достигать 40 - 60 мг на дм2 поверхности листа в час, что вдвое выше, чем для большинства сельскохозяйственных растений. Способность C4-растений концентрировать CO2 в составе C4-кислот является важным адаптивным признаком при водном стрессе, дефиците углекислоты и повышенных температурах, когда растворимость и доступность CO2 понижены. В условиях низкой влагообеспеченности C4-растения продуктивнее C3-видов.
Ю. С. Карпилов и И. А. Тарчевский впервые обнаружили в растениях кукурузы существование пути ассимиляции CO2, принципиально отличающегося от цикла Кальвина [13; 27]. Позднее этот путь был описан в растениях сахарного тростника М. Хэтчем и К. Слэком. Акцептором CO2 в нем является не рибулезобисфосфат (РБФ), а фосфоенолнируват (ФЕП). Основным ферментом, фиксирующим CO2, является фосфо- енолпируваткарбоксилаза (ФЕП-карбоксилаза). Первичным продуктом усвоения углекислого газа является малат или аспартат (C4-цикл дикарбоновых кислот), и только через некоторое время усвоенный углерод из C4-кислот (малата, аспартата) переходит в фосфорилированные сахара (по циклу Кальвина) (рис. 11). К растениям с C4-типом фотосинтеза относятся сахарный тростник, кукуруза, различные виды проса, ряд ксерофитных форм. В этих растениях присутствуют оба цикла фиксации CO2 - цикл Кальвина и цикл Карпилова, Хэтча - Слэка (C4-цикл дикарбоновых кислот). C4-цикл дикарбоновых кислот обнаруживается в хлоропластах с хорошо развитыми гранами (клетки мезофилла), а в крупных хлоропластах, часто не имеющих гран (клетки обкладки проводящих пучков), локализованы ферменты цикла Кальвина. Подобная компартментация углеродного метаболизма позволяет ксерофитам осуществлять фотосинтез даже при закрытых устьицах, так как хлоропласты клеток обкладки используют при этом малат (аспартат) в качестве донора CO2, который раньше образовался в клетках мезофилла листа при фиксации углекислого газа с участием ФЕП-карбоксилазы.
Следовательно, C4-растения могут осуществлять фотосинтез даже при закрытых устьицах, что в жаркое время дня сокращает потери воды за счет транспирации. Кроме того, C4-растения обладают высокой фотосинтетической активностью. У C3-растений даже на свету выделяется из клеток значительное количество CO2 (до 50%) при световом дыхании (фотодыхании). Для формирования 1 г сухого вещества C3-растениям требуется в среднем 450 - 900 г воды. Растения C4-типа, почти не обладающие световым дыханием, производят то же количество сухой массы, используя всего 250 - 350 г воды. C4-тип фотосинтеза протекает с более благоприятным соотношением CO2 и H2O и представляет пример более эффективной экологической адаптации, особенно для тропических областей. В засушливых местах обитания C4-растения имеют преимущества перед C3-растениями.
Суккуленты (роды Crassula, Bryophyllum и др.) также приспособлены осуществлять фотосинтез в условиях засушливого климата. Для них характерен суточный цикл метаболизма C4-кислот с образованием малата ночью. Этот тип фотосинтеза называют САМ-метаболизмом (от английских слов crassulaccan acid metabolism).
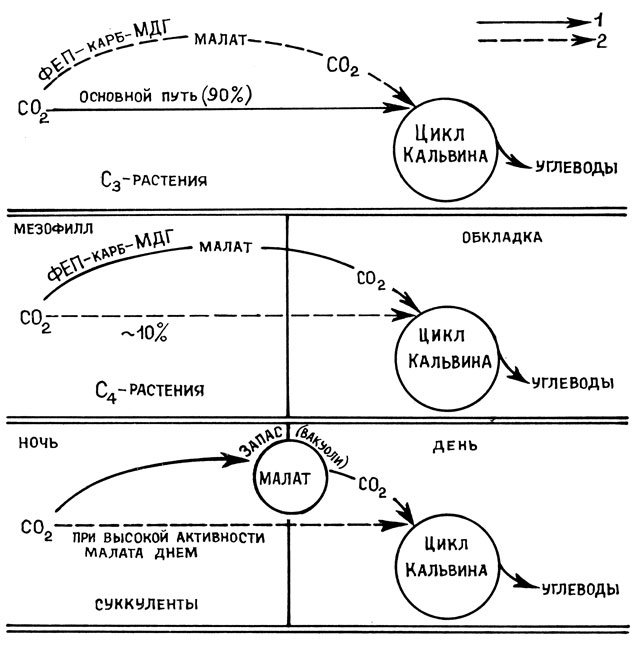
Рис. 11. Сравнительное представление потоков углерода при фиксации CO2 у растений с C3-фотосинтезом (цикл Кальвина), C4-фотосинтезом (путь Хэтча и Слэка), а также у суккулентов (по Kluge, 1971): 1 - основные потоки углерода; 2 - побочные пути потоков углерода
Адаптация к засухе у САМ-растений (очиток едкий, алоэ, заячья капуста) происходит на уровне хлоропластов, которые при водном дефиците обладают способностью оттягивать воду из водоносной паренхимы листа. Устьица у суккулентов днем закрыты, а ночью открываются. В результате растения получают необходимый CO2 ночью, а днем при закрытых устьицах избегают потерь воды. Таким путем суккуленты приспосабливаются к засушливым условиям местообитания. Процессы фотосинтеза у суккулентов и C4-растений во многом сходны. Отличие состоит в том, что у суккулентов процессы фиксации CO2, катализируемые ФЕП-карбоксилазой и РБФ-карбоксилазой, разделены не в пространстве между мезофиллом и клетками обкладки (у C4-растений), а во времени (день и ночь) (рис. 11). Одной из особенностей суккулентов является то, что в ночное время у них синтезируется большое количество яблочной и изолимонной кислот. В течение дня, когда идут процессы фотосинтеза, эти кислоты исчезают. Задача, стоящая перед данными растениями, заключается в том, чтобы ночью накопить CO2, а днем включить ее в состав органических соединений путем фотосинтеза. При темновой фиксации CO2 ФЕП-карбоксилаза выступает у суккулентов в качестве ключевого фермента:
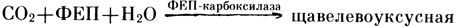
кислота +Фн. Затем щавелевоуксусная кислота с помощью малатдегидрогеназы восстанавливается в малат. Следует отметить, что многие суккуленты накапливают значительное количество малата в цитоплазме и особенно в вакуолях. Малат служит резервным материалом, в котором связывается атмосферный CO2. Днем, когда устьица открыты, малат транспортируется из вакуолей в цитоплазму и там происходит высвобождение из него CO2. Эта реакция катализируется "маликэнзимом":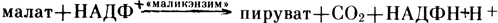 . В результате происходит понижение уровня малата, накопленного ночью. Освобожденный CO2 поступает в хлоропласты и через цикл Кальвина включается в синтез углеводов. Таким образом, у суккулентов работают две фиксирующие CO2 системы: протекающий на свету цикл Кальвина и работающая в темноте система ФЕП-крабоксилаза/малатдегидрогеназа (рис. 11). Следует отметить, что при достаточном количестве воды в клетках некоторые растения с метаболизмом по CAM-типу могут вести себя как C3-растения. При водном дефиците ряд растений с C3-типом фотосинтеза проявляют черты САМ-метаболизма.
. В результате происходит понижение уровня малата, накопленного ночью. Освобожденный CO2 поступает в хлоропласты и через цикл Кальвина включается в синтез углеводов. Таким образом, у суккулентов работают две фиксирующие CO2 системы: протекающий на свету цикл Кальвина и работающая в темноте система ФЕП-крабоксилаза/малатдегидрогеназа (рис. 11). Следует отметить, что при достаточном количестве воды в клетках некоторые растения с метаболизмом по CAM-типу могут вести себя как C3-растения. При водном дефиците ряд растений с C3-типом фотосинтеза проявляют черты САМ-метаболизма.
Фотосинтез является сложным, многоступенчатым процессом и осуществляется с участием различных ферментных систем. Засуха может оказывать влияние на отдельные звенья этого процесса.
Е. Ф. Вотчал с сотрудниками, при исследовании влияния недостатка влаги в почве на фотосинтез, установил, что в жаркую погоду наблюдается депрессия данного процесса, вызванная нарушением водного режима растений. Результаты, полученные А. С. Оканенко с сотрудниками, свидетельствуют о том, что снижение влажности листьев с 83 до 81% не отражалось на интенсивности фотосинтеза, а при дальнейшем понижении содержания воды до 79% интенсивность фотосинтеза падала до 76% [5].
Многие исследователи отмечают двухфазную реакцию фотосинтеза на обезвоживание (рис. 10). Полагают, что в начале обезвоживания в общем снижении фотосинтеза главную роль играет степень открытости устьиц, а при более длительном воздействии засухи приобретают значение нарушения фотохимических реакций и других ферментативных процессов.
В условиях водного дефицита интегральные белки хлоропластных мембран подвергаются меньшему воздействию, чем белки стромы (12].
Электронно-микроскопические наблюдения показывают, что структура фотосинтетических мембран многих растений (кукурузы, сорго, пшеницы) весьма устойчива к условиям обезвоживания. По-видимому, структурная прочность мембран является важным фактором в способности растений противостоять водному стрессу. В условиях значительного водного стресса резкое повышение внутриклеточной активности гидролитических ферментов в листьях хлопчатника приводит к разрушению структуры митохондрий, хлоропластов, пластидных рибосом. В частности, в митохондриях частично обезвоженных растений хлопчатника выявлена высокая активность кислой липазы [35].
Многие исследователи отмечают лабильность мембранных систем хлоропластов, которые после непродолжительного обезвоживания и последующей гидратации быстро восстанавливают свою ультраструктуру и функциональную активность. Состояние мембранных компонентов и хлорофилл-бел нового комплекса в хлоропластах в значительной степени зависит от устойчивости растений к засухе. У относительно засухоустойчивых растений (персик) ультраструктура хлоропластов при непродолжительном (4 часа) подсушивании листьев не меняется, тогда как у менее устойчивых (яблони, груши) - в хлоропластах обнаруживается разбухание краевых дисков гран, дезорганизация мембран и другие изменения. Связь пигментов с липопротеидным комплексом мембран у засухоустойчивых растений более прочная по сравнению с менее устойчивыми растительными формами.
Исследования, проведенные С. А. Алиевой с сотрудниками на растениях нута таджикского (ксерофит) и киевского (мезофит), показали, что в условиях низкой водообеспеченности (влажность почвы 30% от полной влагоемкости) у данных организмов уже на 15-й день наблюдаются первые признаки изменения внутренней структуры хлоропластов (набухание ламелл и увеличение межламеллярных пространств). При чем, у мезофитных форм эти изменения были более выраженными. Нарушение структуры хлоропластов у мезофитов вследствие недостатка влаги сопровождалось снижением на 25 - 50% интенсивности фотосинтеза и уровня фотофосфорилирования. У ксерофитных форм, у которых структурные изменения хлоропластов были незначительными, и фотофосфорилирование, и фотосинтез практически оставались неизменными в течение 30 дней дефицита влаги в почве [5].
А. Л. Курсанов отмечает, что недостаточная влагообеспеченность растений приводит к снижению скорости передвижения ассимилятов. В случае прогрессирующей почвенной засухи у пшеницы снижается интенсивность фотосинтеза, ослабевает поступление ассимилятов в окончания проводящих путей и уменьшается масса транспортируемых ассимилятов. У растений кукурузы транспорт продуктов фотосинтеза подавляется под действием водного дефицита раньше и в большей степени, чем сам процесс фотосинтеза. При подвядании сахарного тростника загрузка ассимилятами флоэмных окончаний также испытывает более сильное торможение, чем фотосинтез [16].
Следовательно, фотосинтез в процессе обезвоживания мезофитных растений изменяется следующим образом: при неглубокой и непродолжительной засухе его интенсивность несколько возрастает, а при сильной и продолжительной - резко снижается (рис. 10). При значительном водном дефиците подавление фотосинтеза вызывается не только повреждением самих хлоропластов, которые в определенной степени устойчивы к обезвоживанию, но и задержкой оттока ассимилятов из листьев, связанной с уменьшением их потребления вследствие торможения ростовых процессов. В условиях сильной и продолжительной засухи продукты фотосинтеза потребляются медленнее, чем они вырабатываются, что приводит к "перекорму" листьев. Накопление избыточного количества продуктов ассимиляции CO2 приводит к ингибированию процесса фотосинтеза.
Транспорт ассимилятов в значительной степени зависит от обеспеченности проводящих тканей АТФ. При обезвоживании мезофитных растений процесс сопряжения дыхания и окислительного фосфорилирования имеет двухфазный характер. В начальный период обезвоживания усиливается сопряжение дыхания и аккумуляции энергии, а при значительной потере воды наступает разобщение этих процессов. Высокий уровень АТФ и усиление интенсивности фотосинтеза (эффект Бриллиант) на первом этапе обезвоживания обеспечивают усиление транспорта ассимилятов у пшеницы. При дальнейшей потере воды клеткой наблюдается падение фотосинтеза, транспорта ассимилятов и разобщение окисления и фосфорилирования, что и приводит к подавлению ростовых и формообразовательных процессов. В результате в условиях длительной и повторяющейся засухи общая продуктивность растительных организмов снижается.
В. Н. Жолкевич детально исследовал энергетику дыхания растений в условиях водного дефицита. Он показал, что дыхание обнаруживает большую устойчивость к обезвоживанию, чем фотосинтез. Характер изменения интенсивности дыхания определяется целым комплексом сопутствующих факторов: скоростью, глубиной и продолжительностью обезвоживания; видом засухи; физиологическим состоянием и биологическими особенностями растений. У большинства засухоустойчивых растений даже при значительном водном дефиците дыхание либо активируется незначительно, либо не изменяется. У незасухоустойчивых видов уже при сравнительно слабом водном дефиците дыхание, как правило, заметно возрастает, а при сильном обезвоживании наблюдается резкое снижение интенсивности данного процесса [9].
При этом в условиях продолжительной и сильной засухи или быстрого наступления обезвоживания клеток дыхание теряет свою эффективность. Уровень окислительных процессов в этих условиях не соответствует синтетическим, а процессы, непосредственно связанные с затратами энергии (поддержание структуры протоплазмы, биосинтезы, транспорт ассимилятов, рост и др.), значительно тормозятся. По- видимому, в условиях продолжительной и сильной засухи энергия дыхания в меньшей степени тратится на метаболические процессы и в основном выделяется в окружающую среду в виде тепла. Причем расход энергии дыхания в условиях засухи на рост, поглощение ионов и поддержание физиологического состояния структур различен. В частности, дыхание, направленное на поддержание физиологического состояния структур, снижается при водном стрессе в меньшей степени по сравнению с дыханием, направленным на рост и поглощение ионов.
Н. А. Сатарова и другие считают, что наиболее чувствительным показателем устойчивости растений к обезвоживанию является интенсивность синтеза белка. В исследованиях, проведенных с интактными растениями картофеля, бобов, кукурузы, фасоли, показано, что при водном стрессе снижается включение 15N в белки листьев, корней. Степень замедления синтеза белка зависит от глубины обезвоживания и устойчивости растений. При поливе растений синтез белка в листьях через сутки начинает восстанавливаться, а через 5 суток включение 15N достигает уровня растений, не подвергавшихся обезвоживанию. У растений кукурузы, закаленных к засухе перед посевом, а также у фасоли и картофеля после теплового закаливания наблюдается увеличение интенсивности синтеза белка при регидратации по сравнению с незакаленными растениями. При обработке растений растворами аденина, кинетина, сернокислого цинка наблюдается даже усиление синтеза при их обезвоживании и повышение устойчивости к засухе и высоким температурам [24].
Таким образом, для мезофитных растений синтез белка, по-видимому, необходим для процессов репарации внутриклеточных структур и адаптации их к обезвоживанию.
П. А. Генкелем было показано, что у кукурузы, бобов, фасоли и разных сортов пшеницы под влиянием засухи и повышенных температур снижается содержание суммарной РНК и падает уровень общего и белкового азота, а также снижается включение 15N в белки, что свидетельствует об изменениях в белоксинтезирующей системе. Кроме того, при обезвоживании клеток значительно увеличивается рибонуклеазная активность. Цитоплазматической рибонуклеазе принадлежит важная регуляторная роль в синтезе белка: она разрушает все виды РНК, и в первую очередь информационную. При наличии водного дефицита повышается суммарная активность рибонуклеазы, в результате чего нарушается сбалансированность белоксинтезирующей системы и снижается уровень белка в клетках [5].
Е. К. Творус и др., рассматривая причины усиления PHКазной активности при обезвоживании, отмечают, что это, возможно, происходит в результате освобождения данного белка из липопротеидного комплекса и из клеточных структур (хлороиластов, митохондрий, ядер), а также за счет высвобождения фермента из неактивных комплексов и синтеза его de novo 1291.
При водном дефиците иод влиянием РНКазы разрушение информационной РНК ведет к деградации полирибосом. Из компонентов белоксинтезирующего аппарата вегетирующих мезофитных растений наиболее чувствительны к обезвоживанию полирибосомы. При значительном водном дефиците в растительных клетках уменьшается количество поли рибосом и увеличивается количество моносом, а это приводит к снижению активности белоксинтезирующей системы.
В растительных клетках хроматин может существовать в двух состояниях: в виде сильно уплотненного (конденсированного) - гетерохроматина и более диспергированного (диффузного) - эухроматина. Достаточное количество данных позволяет считать, что гены, локализованные в гетерохроматиновых участках хромосом, лишены активности. При нарастании водного дефицита обнаруживается значительное увеличение доли гетерохроматина (прочносвязанной ДНК), что свидетельствует о нарушении трансляции генетической информации на этапе синтеза матричных РНК.
Таким образом, причинами нарушения процесса синтеза белков при водном дефиците является повышение рибонуклеазной активности, распад полирибосом и уменьшение способности передачи информации из ядра в клетку.
Установлено также, что при действии засухи соотношение между синтезом и гидролизом веществ сдвигается в сторону гидролиза, а падение содержания белка ниже критического уровня ведет к повреждению и гибели растений. Работами К. Мотеса было показано, что при обезвоживании клеток происходит распад белков и образование большого количества аммиака. Избыточный аммиак, который может оказывать повреждающее действие на клетку, связывается аминокислотами с образованием амидов или органическими кислотами с образованием аммонийных солей. В частности, накопление аланина является приспособительной реакцией, направленной на снижение уровня аммиака в клетках засухоустойчивых растений [27; 28].
Первые сообщения о повышении содержания пролина в растениях в условиях засоления и водного дефицита относятся к 1960-м г., когда была высказана гипотеза о действии пролина как осморегулятора. В дальнейшем осморегуляторное действие пролина стали рассматривать как один из факторов резистентности растений и зеленых водорослей к солям и недостатку воды. Осморегуляция - действенное и быстрое средство, благодаря которому не допускается осмотическое высушивание клеточного содержимого. Это достигается избыточным поглощением ионов и аккумуляцией в клетках пролина. Однако высокие концентрации солей угнетают активность большинства ферментов. Осморегуляция, основанная на увеличении водного потенциала путем усиленного накопления осмотически активных органических веществ, в частности пролина, способствует удержанию воды и практически безвредна для большинства ферментов. Пролин - компонент осморегуляторного механизма. Кроме того, он обладает и протекторными свойствами. Взаимодействие агрегатов молекул пролина с белками повышает растворимость последних и защищает их от денатурации при водном дефиците. Низкомолекулярные протекторы, тот же пролин, способны изменять состояние воды в биологических мембранах таким образом, что утраченная при дегидратации оводненность мембран восстанавливается. Существуют пролинаккумулирующие виды, у которых содержание данной аминокислоты может повышаться при стрессе в 100 и более раз (ячмень, шпинат, арахис, хлопчатник, сведа, персик, C4-травы). У других растений (подсолнечник, многие суккуленты) вклад пролина в осморегуляцию минимален. Такая неодинаковая способность к аккумуляции пролина, очевидно, является генетически детерминированной нормой реакции на стресс у различных видов растений. Сверхнродукция пролина в условиях водного стресса является одним из механизмов метаболической адаптации растительных организмов.
Дефицит влаги вызывает изменение сбалансированного уровня фитогормонов в клетках мезофитных растений: увеличивается содержание абсцизовой кислоты и уменьшается содержание стимуляторов роста. При обезвоживании клеток антагонизм между АБК и цитокинином проявляется на уровне процессов, связанных с мембранами и приводящих к открыванию и закрыванию устьиц.
Наблюдаются различия в особенностях гормональной регуляции роста у суккулентов и мезофитов в условиях засухи. Установлено, что завядание обрезанных побегов опунции (суккулент) в течение 24 дней сопровождается увеличением содержания ауксинов и некоторым снижением ингибиторов роста. Сходная закономерность обнаруживается в изменении эндогенных регуляторов роста также у опунции, подвергающейся длительной (69 дней) почвенной засухе в период формирования бутонов и цветов. Способность поддерживать значительный уровень веществ, активирующих рост, позволяет данным растениям возобновлять ростовые процессы и способствует быстрому образованию корней с прекращением засухи.
У мезофитов при обезвоживании происходит снижение содержания стимуляторов роста и накопление ингибиторов. При этом у мезофитов, обладающих значительной засухоустойчивостью (персик, а также закаленные растения абрикоса), на начальных этапах засухи незначительно изменяется уровень стимуляторов и накапливается меньше ингибиторов роста.
Следовательно, соотношение стимуляторов и ингибиторов у мезофитов сдвигается в сторону накопления ингибиторов, которые и определяют прекращение ростовых процессов этих растений в условиях засухи. У суккулентов соотношение между фитогормонами сдвигается в сторону некоторого увеличения содержания активаторов и снижения уровня ингибиторных веществ, и в результате в условиях водного стресса скорость роста у них и практически не изменяется.
Борьба с засухой и повышение устойчивости растений. Наиболее радикальное средство борьбы с засухой - применение орошения, которое позволяет увеличить урожай сельскохозяйственных культур в 2 - 3 раза [21; 22]. Кроме того, целесообразно проводить следующие мероприятия:
1) Использовать для посева наиболее засухоустойчивые сорта и высевать их в наилучшие сроки.
2) Высевать те или иные культуры по чистому пару, содержащему, как правило, больше воды в почве по сравнению с почвами после предшественников.
3) Вносить в почву органические и минеральные удобрения, которые обеспечивают более экономное расходование воды.
4) Вести активную борьбу с сорной растительностью, болезнями и вредителями.
5) Проводить предпосевное закаливание растений.
Метод закаливания предложен П. А. Генкелем и заключается в замачивании семян в чистой воде (соотношение 1:2) до полного их набухания и наклевывания с последующим подсушиванием при температуре 20 - 22° в течение 30 часов до воздушно-сухого состояния. Растения, полученные из закаленных семян, обнаруживают глубокую перестройку обмена веществ: у них повышается вязкость протоплазмы, увеличивается интенсивность фотосинтеза, активируются дыхательные ферменты. Митохондрии у закаленных растений более устойчивы к перегреву и быстрее восстанавливают свою фосфорилирующую активность при поливе после перенесенной засухи. У закаленных растений менее активна рибонуклеаза и в них интенсивнее протекает синтез белка. Закаливание стабилизирует структуру мембран и увеличивает устойчивость растений одновременно к перегреву и обезвоживанию.
6) Проводить обработку семян аденином, сернокислым цинком, кинети ном и картолином.
Эти химические вещества способствуют частичному подавлению активности рибонуклеазы и стабилизируют содержание РНК, белка и биосинтез белка в условиях засухи. Из работ О. Н. Кулаевой следует, что цитокинины задерживают деградацию структурных элементов в стрессовых условиях, а также защищают от разрушения белки, нуклеиновые кислоты и липиды. Кроме того, цитокинины стимулируют деление клеток и задерживают старение. Все это способствует процессу закаливания и повышению устойчивости растений [13].
В настоящее время ведется поиск аналогов цитокининов, обладающих большей способностью повышать устойчивость растений к неблагоприятным условиям среды. К таким соединениям относится, например, картолин, который по сравнению с цитокининами лучше защищает растения от повреждения засухой [31; 29].
Обработка растений ячменя раствором картолина за несколько дней до наступления засухи предохраняла от распада полирибосомы и нормализовала активность РНК-полимеразы в клетках листьев в условиях недостаточного водоснабжения. Кроме того, под влиянием картолина увеличивался урожай обработанных растений по сравнению с необработанными, а также повышалась устойчивость клеток листа к высоким температурам и обезвоживанию.
7) Использовать ретарданты, в частности хлорхолинхлорид (ССС).
Под влиянием ССС снижается интенсивность роста стеблей злаков и они становятся менее требовательными к воде и более устойчивыми к завяданию. Обработанные ССС зерновые злаки приобретают ряд свойств, характерных для засухоустойчивых растений: они низкорослы, имеют мощно развитую, глубоко проникающую корневую систему, слабее транспирируют, имеют повышенную оводненность тканей листьев и стебля и пониженную проницаемость протоплазмы для электролитов. Эти свойства проявляются как при опрыскивании растений ретардантом, так и при предпосевной обработке семян. Растения из семян, обработанных ретардантом, не теряли способности к фотосинтезу в жестких метеорологических условиях, когда температура на поверхности почвы достигала +55°С, температура воздуха в посевах пшеницы +35°С. При этом предпосевная обработка семян ССС заметно повышает устойчивость пшеницы как к засухе, так и к зимним холодам (положительная сопряженная устойчивость).
Таким образом, исследование особенностей метаболизма растений, различающихся по устойчивости к засухе, показало, что защитно-приспособительные реакции носят комплексный характер и затрагивают различные стороны обмена веществ. Устойчивые растения характеризуются незначительными нарушениями структуры и функции и более быстрым их восстановлением в оптимальных условиях водоснабжения. У неустойчивых растений в условиях значительного водного дефицита происходят в основном необратимые нарушения физиолого-биохимических процессов. Адаптация к засухе вызывает у растений стимуляцию ряда метаболических и физиологических процессов, что позволяет повышать продуктивность и получать в условиях засухи более устойчивые урожаи.
На растения в естественных условиях их обитания оказывает влияние не один фактор, а целая система сопутствующих условий, и в каждом конкретном случае их соотношение различно. Интенсивность роста и развития растений во многом зависит от напряженности экологических факторов. При этом каждому этапу органогенеза соответствует свой экологический оптимум и в зависимости от того, насколько условия произрастания соответствуют последнему, изменяется жизненный ритм и продуктивность растений.
Отбор растений по их сельскохозяйственной или декоративной ценности нередко приводит к ослаблению адаптивной приспособляемости организмов к факторам окружающей среды. Многие культурные растения утратили в связи с этим способность к независимому существованию и полностью зависят от антропогенного фактора, который обеспечивает им благоприятные условия для поддержания нормального существования. В сельскохозяйственной практике издавна применяются различные приемы воздействия на семена и молодые проростки растений физическими, химическими факторами и физиологически активными веществами, направленными на улучшение их посевных и урожайных качеств, повышение устойчивости растений к болезням и неблагоприятным факторам окружающей среды.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'