Влияние температуры
Реакции на температуру, так же как и на свет, у растений могут быть и качественными и количественными. Скорости почти всех химических процессов в растении с повышением температуры градуально (т. е. плавно) возрастают, достигают некоторого максимума, а затем снижаются. В отличие от этого многие онтогенетические процессы, например прорастание семян и прерывание покоя почек, часто регулируются по принципу "всё или ничего". В последних случаях для низкотемпературной индукции требуется непрерывное воздействие в течение какого-то минимального периода; это напоминает фотопериодическую индукцию, при которой тоже необходимы совершенно определенные периоды темноты.
Градуальные реакции
Скорость большинства химических процессов неуклонно возрастает с повышением температуры. Температурный коэффициент Q10 показывает, во сколько раз увеличивается скорость реакции при повышении температуры на 10° по сравнению с первоначальной:
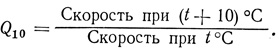
Q10 для большинства химических (и в том числе ферментативных) реакций равно по меньшей мере 2, тогда как для физических процессов, таких как диффузия или фотохимические реакции, Q10 составляет около 1,1 или 1,2. У растений, выращиваемых при температурах от 0 до 30 °С, с повышением температуры постепенно увеличивается скорость вытягивания и Q10 равно около 2,0 или больше (рис. 12.21). По неизвестным нам причинам температурные оптимумы для разных растений очень сильно различаются, и это указывает на то, что какой-то фундаментальный биохимический процесс у них обладает различной чувствительностью к температуре. В зоне выше оптимума абсолютная скорость роста у всех растений снижается, иногда очень резко. Для большей части растений этот оптимум лежит в области от 28 до 32 °С.
Мы не знаем, почему большинство растений повреждается температурами выше примерно 30 °С, хотя ферменты или органеллы, выделенные из растений, при таких температурах обычно не повреждаются. Одно из возможных объяснений состоит в том, что мембраны клеток или их органелл чувствительны к изменениям температуры из-за плавления или затвердевания жирных кислот в фосфолипидах. Известно, что растения при более низких температурах синтезируют больше ненасыщенных жиров с соответственно более низкой температурой плавления.
Обратное явление наблюдается при высоких температурах (табл. 12.1). Другая возможность состоит в том, что при повышенных температурах какие-то материалы, необходимые для роста, очень быстро разрушаются или не образуются в нужном количестве. У многих организмов имеются гены "чувствительности к температуре". У плесневого гриба Neurospora ген, ответственный за образование витамина B2 (рибофлавина), хорошо функционирует при низких температурах, но не действует нормально при более высоких. Поэтому при 35 °С гриб должен непременно получать рибофлавин извне, тогда как при. 25 °С он может расти и в отсутствие этого вещества. Подобным образом дело могло бы обстоять с другими витаминами, аминокислотами или гормонами у высших растений. Если это так, то, зная химическую основу индуцированного температурой замедления роста при супраоптимальных температурах, можно было бы улучшить рост путем внесения нужных веществ.
Морфогенетические эффекты
Очень малая скорость химических реакций в растении при низких температурах обеспечивает координацию изменений роста с климатическими изменениями. Кроме того, температура влияет на многие процессы, чувствительные к фотопериоду, изменяя критическую длину темного периода, хотя механизм этого явления не выяснен. Так как наличие Фдк ночью могло бы нарушать индукцию цветения у растения короткого дня, можно - было бы ожидать, что высокие ночные температуры, ускоряющие разрушение фитохрома и превращение Фдк в Фк, будут благоприятствовать индукции цветения у таких растений. На самом же деле обычно наблюдается противоположный эффект. Некоторые растения можно заставить цвести с помощью длинных темных ночей или низких ночных температур. Возможно, что каждое из этих воздействий способно активировать какой-то процесс, ведущий к синтезу флоригена.
К наиболее драматичным эффектам низкой температуры относятся прорастание семян, прерывание покоя почек (рис. 12.22) и подготовка к закладке цветочных бугорков. Низкотемпературное воздействие, способствующее прорастанию семян, называют стратификацией, а облегчающее инициацию цветения - яровизацией. Первое препятствует преждевременному прорастанию, а второе обеспечивает двулетний тип развития. Так как оптимальные температуры, необходимые для прорастания и яровизации, для разных растений различны, температура является важным фактором, влияющим на географическое распространение растений и регулирующим время цветения и плодообразования.

Рис. 12.22. А. Зимняя веточка конского каштана (Aesculus hippocastanum) со спящими почками и подковообразными рубчиками, оставшимися после опадения листьев в предыдущем году. После прерывания покоя холодным зимним периодом почки с наступлением весны лопаются. Б-Г. Постепенное распускание почек в течение примерно 10 дней с образованием новых листьев и цветочных почек
Значительная часть сведений о яровизации была получена в исследованиях, проведенных на однолетних и двулетних линиях ржи (Secale cereale). Однолетняя линия заканчивает свой репродуктивный цикл за один вегетационный период, а двулетней для цветения необходима перезимовка в поле. Двулетник в первый год образует только вегетативные органы; цветочные бугорки не могут быть заложены до тех пор, пока растение не подвергнется длительному воздействию низких температур. Только после такого воздействия оно способно реагировать на надлежащий фотопериодический стимул. Эта потребность в низких температурах может быть удовлетворена в любую фазу онтогенеза после прорастания семян. Например, если семени двулетника позволить поглотить некоторое количество воды для начала прорастания, а затем около шести недель держать при низких температурах (примерно 2-5°С), растение будет вести себя так, как будто оно пережило холодную зиму после года роста, и зацветет при весеннем посеве, если будет соответствующий фотопериод (рис. 12.23). Так как различия между однолетними и двулетними расами у большинства растений, по-видимому, зависят от одного гена, низкая температура для двулетней формы, вероятно, служит заменой какому-то генетически контролируемому биохимическому событию, происходящему у однолетней формы без воздействия холода.
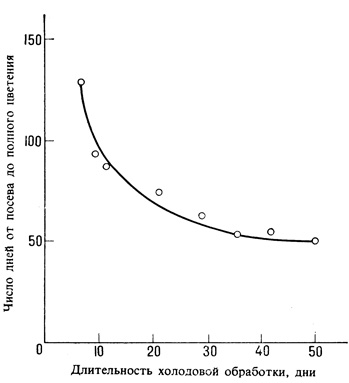
Рис. 12.23. Некоторые двулетники без холодовой обработки не зацветают совсем. Рожь дает количественную реакцию: с увеличением продолжительности холодовой обработки период от посева до цветения сокращается. (Purvis, Gregory. 1937. Ann. Bot. N. S, 1, 569-592.)
Холодовой стимул воспринимается верхушкой побега, тканью, в которой происходит и образование цветочных почек. Поэтому эффекту такого стимула в отличие от флоригена, индуцируемого фотопериодом, нет необходимости перемещаться по растению. Большинство растений, требующих яровизации, являются длиннодневными, и им нужна фотопериодическая индукция через некоторое время после прорастания. Это позволяет предполагать, что холод вызывает какое-то локальное изменение клеток, которое дает им возможность реагировать на флориген. Изменения, вызванные низкой температурой, у большинства растений после индукции сохраняются неопределенно долгое время. Например, яровизированное состояние воспроизводится при тысячах клеточных делений в апикальных меристемах; таким образом, яровизации какой-либо почки достаточно для передачи ее эффекта всем клеткам, возникающим из этой почки.
У некоторых двулетников воздействие гиббереллином на растение, не подвергнутое яровизации, видимо, заменяет низкую температуру, вызывая при благоприятном фотопериоде быструю закладку цветочных бугорков (рис. 9.21). Но так как гиббереллины не всегда могут заменять холодовую обработку, кажется маловероятным, что эффект яровизации всецело обусловлен увеличением синтеза гиббереллинов. Согласно другому предположению, яровизация "дерепрессирует" особые гены в обработанных холодом клетках - снимает блокаду, препятствовавшую специфическим процессам транскрипции и трансляции. Это привлекательная гипотеза, так как и при стратификации, и при яровизации усиливается синтез многих ферментов.
Прорастание семян
Температурные потребности для прорастания семян значительно варьируют в зависимости от вида. Например, для однолетнего Delphinium оптимальна температура 15°С, а для дыни - от 30 до 40 °С. Однако многие семена остаются в состоянии покоя даже при увлажнении и подходящей для прорастания температуре. Иногда такой покой можно прервать светом (гл. 11), путем преинкубации при определенной температуре, увеличения количества O2, вымывания ингибиторов, физического разрушения оболочек семени или плода. Семена растений умеренной зоны часто требуют низкотемпературной стратификации перед инкубацией при более высоких температурах; это задерживает прорастание до весны, предотвращая возможное повреждение нежных проростков морозами. С другой стороны, семена некоторых растений, например белого дуба, прорастают сразу после падения на землю и быстро формируют глубоко идущую корневую систему.
Сухие семена не реагируют на стратификацию, и перед воздействием низкой температурой им обычно дают полностью набухнуть. Предполагают, что при обработке происходят какие-то метаболические трансформации, которые проявляются в изменении интенсивности дыхания, путей транслокации, содержания ферментов и даже скорости деления и растяжения клеток. Один очень важный комплекс изменений касается баланса гормонов: уровни АБК обычно снижаются, в то время как содержание гиббереллинов (в одних семенах) или цитокининов (в других) возрастает. АБК, как правило, связана с покоем, а гиббереллины и цитокинины - с процессами синтеза и роста. Стратификация, таким образом, приводит к "послеуборочному дозреванию" и тем самым биохимически подготавливает к последующему прорастанию при более высокой температуре.
Семенам некоторых растений, главным образом пустынных, перед прорастанием необходима стратификация при высоких температурах. В других случаях существует взаимосвязь между условиями температуры и освещения. Например, семена щирицы Amaranthus retroflexus прорастают в темноте, если держать их несколько дней при 20 °С, а затем перенести на 35 °С. Но если исключить низкие температуры, то для прорастания необходим свет. Это, по-видимому, указывает на потребность в Фдк. Вспомните, что значительная часть фитохрома в семенах находится в форме Фдк, который после увлажнения семян может снова превращаться в Фк. Так как обращение Фдк у Amaratithus при 20 °С идет очень медленно, а при 25 °С - в четыре раза быстрее, требующие Фдк процессы, которые ведут к прорастанию, могут происходить в период охлаждения. Таким образом, некоторые специфические потребности в температуре и свете для прорастания мы можем поставить в связь с Фдк, а другие - с гормонами. Каким бы ни был регуляторный механизм, он должен в конечном итоге влиять на активность генов и сам должен находиться под ее влиянием.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'