Растительность влажного тропического леса
Строение и структура. Дать обобщенное описание структуры влажного тропического леса практически невозможно: это сложнейшее растительное сообщество обнаруживает такое разнообразие типов, что их не в состоянии отразить и самые подробные описания. Еще несколько десятилетий назад полагали, что влажный лес - это всегда непроходимые заросли деревьев, кустарников, наземных трав, лиан и эпифитов, поскольку в основном судили о нем по описаниям горных влажных лесов. Лишь сравнительно недавно стало известно, что в некоторых влажных тропических лесах из-за плотной сомкнутости крон высоких деревьев солнечный свет почти не доходит до почвы, поэтому подрост здесь скудный, и через такие леса можно пройти почти беспрепятственно.
Принято особо подчеркивать видовое разнообразие влажного тропического леса. Часто отмечают, что в нем едва ли найдешь два экземпляра деревьев одного и того же вида. Это - явное преувеличение, но вместе с тем отнюдь нередко на площади в 1 гектар можно встретить 50-100 видов деревьев.
Но имеются и относительно бедные видами, "монотонные" влажные леса. К ним относятся, например, особые леса, состоящие в основном из деревьев семейства двукрылоплодниковых (Dipterocarpaceae), растущие в очень богатых осадками областях Индонезии. Их существование свидетельствует о том, что в этих областях стадия оптимального развития влажных тропических лесов уже пройдена. Крайнее обилие осадков затрудняет аэрацию почвы, в результате произошел отбор растений, приспособившихся к обитанию в таких местах. Сходные условия существования можно встретить также в некоторых сырых районах Южной Америки и бассейна Конго.
Господствующий компонент влажного тропического леса - деревья разного внешнего вида и разной высоты; они составляют около 70% всех встречающихся здесь видов высших растений. Различают три яруса деревьев - верхний, средний и нижний, которые, правда, редко выражены четко. Верхний ярус представлен отдельными гигантскими деревьями; их высота, как правило, достигает 50-60 м, а кроны развиваются над кронами деревьев ниже расположенных ярусов. Кроны таких деревьев не смыкаются, во многих случаях эти деревья рассеяны в виде отдельных экземпляров, кажущихся переростками. Напротив, кроны деревьев среднего яруса, имеющих высоту 20-30 м, обычно образуют сомкнутый полог. Из-за взаимовлияния соседних деревьев их кроны бывают не столь широкими, как у деревьев верхнего яруса. Степень развития нижнего древесного яруса зависит от освещенности. Его составляют деревья, достигающие в среднем примерно 10-метровой высоты. Встречающимся в разных ярусах леса лианам и эпифитам будет посвящен особый раздел книги (стр. 100-101).
Часто имеются также ярус кустарников и один- два яруса травянистых растений, их составляют представители видов, способных развиваться при минимальной освещенности. Поскольку влажность окружающего воздуха постоянно высока, устьица этих растений остаются открытыми в течение всего дня и растениям не грозит увядание. Таким образом, они постоянно ассимилируют.
По интенсивности и характеру роста деревья влажного тропического леса можно разделить на три группы. Первую составляют виды, представители которых быстро растут, но недолго живут; они первыми развиваются там, где в лесу либо естественным путем, либо в результате деятельности человека образуются осветленные участки. Эти светолюбивые растения прекращают свой рост примерно лет через 20 и уступают место другим видам. К числу таких растений относятся, например, южноамериканские бальзовое дерево (Ochroma lagopus) и многочисленные мирмекофильные виды цекропии (Cecropia), африканский вид Musanga cecropioides и произрастающие в тропической Азии представители семейства молочайных, относящиеся к роду Macaranga.
Вторая группа охватывает виды, представители которых на ранних стадиях развития также растут быстро, но их рост в высоту продолжается дольше, и по его окончании они способны жить еще очень долго, вероятно, не одно столетие. Это наиболее характерные деревья верхнего яруса, кроны которых обычно не затенены. В их число входят многие хозяйственно важные деревья, древесину которых принято называть "красным деревом", например виды, относящиеся к родам Swietenia (тропическая Америка), Khaya и Entandrophragma (тропическая Африка).

Профили тропических влажных лесов: на низменности (слева) и в горах (справа)
Наконец, третья группа включает в себя представителей теневыносливых видов, растущих медленно и долгоживущих. Их древесина обычно очень тяжелая и твердая, обрабатывать ее трудно, и поэтому она не находит столь широкого применения, как древесина деревьев второй группы. Тем не менее к третьей группе относятся виды, дающие благородную древесину, в частности Tieghemella heckelii или Aucoumea klainiana, древесина которого используется в качестве заменителя красного дерева.
Для большинства деревьев характерны прямые, колоннобразные стволы, которые часто, не ветвясь, поднимаются более чем на 30-метровую высоту. Только там у отдельно стоящих гигантских деревьев развивается раскидистая крона, тогда как в нижних ярусах, как уже упоминалось, деревья из-за тесного их расположения образуют лишь узкие кроны.
У некоторых видов деревьев около оснований стволов образуются досковидные корни (см. рисунок), порой достигающие высоты до 8 м. Они придают деревьям большую устойчивость, поскольку корневые системы, развивающиеся неглубоко, не обеспечивают достаточно прочного закрепления этих огромных растений. Образование досковидных корней обусловлено генетически. У представителей одних семейств, например у Моrасеае (тутовых), Mimosaceae (мимозовых), Sterculiaceae, Bombacaceae, Meliaceae, Bignoniaceae, Combretaceae, они встречаются довольно часто, а у других, например Sapindaceae, Apocynaceae, Sapotaceae, их вовсе нет.
Деревья с досковидными корнями чаще всего растут на сырых почвах. Возможно, развитие досковидных корней связано с характерной для таких почв плохой аэрацией, препятствующей вторичному приросту древесины на внутренних сторонах боковых корней (она образуется только с наружных их сторон). Во всяком случае, у деревьев, растущих на пропускающих влагу и хорошо аэрированных почвах горных дождевых тропических лесов, досковидных корней нет.
Для деревьев других видов характерны ходульные корни; они образуются выше основания ствола как придаточные и особенно часто встречаются у деревьев нижнего яруса, также растущих преимущественно в сырых местообитаниях.
Различия в микроклимате, свойственные разным ярусам влажного тропического леса, отражаются и на строении листьев. В то время как деревья верхних ярусов обычно имеют эллиптические или ланцетные в очертаниях, гладкие и плотные кожистые листья типа листьев лавра (см. рисунок на стр. 112), способные переносить в течение суток чередование сухих и влажных периодов времени, листья деревьев нижнего яруса обнаруживают признаки, свидетельствующие об интенсивной транспирации и быстром удалении влаги с их поверхности. Они обычно более крупные; их пластинки имеют особые заострения, на которых собирается вода, а затем падает с них каплями, поэтому на поверхности листа нет водной пленки, которая препятствовала бы транспирации.
На смену листвы у деревьев влажных тропических лесов не влияют внешние факторы, в частности засуха или холод, хотя и здесь можно заменить известную периодичность, варьирующую у разных видов. Кроме того, проявляется некоторая самостоятельность отдельных побегов или ветвей, поэтому безлистным бывает не все дерево сразу, а лишь часть его.

Досковидные корни одного из деревьев влажного тропического леса
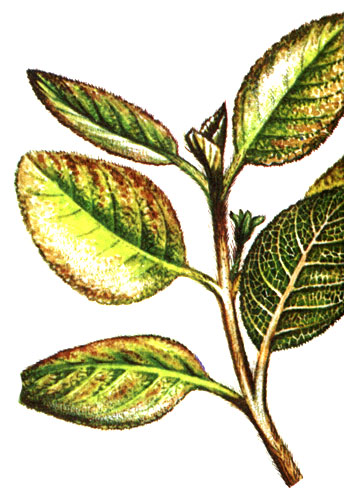
Особенности климата влажного тропического леса сказываются также на развитии листвы. Поскольку здесь нет необходимости защищать точки роста от холода или засухи, как в областях с умеренным климатом, почки выражены сравнительно слабо и не окружены почечными чешуями. При развитии новых побегов у многих деревьев влажного тропического леса наблюдается "поникание" листьев, что вызвано исключительно быстрым увеличением их поверхности. Из-за того что механические ткани не формируются столь же быстро, молодые черешки сначала, словно подвядшие, свисают вниз, листва как бы поникает. Образование зеленого пигмента - хлорофилла - при этом также может замедляться, и молодые листья оказываются беловатыми или - что обусловлено содержанием пигмента антоциана - красноватыми (см. рисунок вверху).

'Поникание' молодых листьев у шоколадного дерева (Theobroma cacao)
Следующая особенность некоторых деревьев влажных тропических лесов - каулифлория, то есть образование цветков на стволах и безлистных участках ветвей. Поскольку это явление наблюдается прежде всего у деревьев нижнего яруса леса, ученые трактуют его как приспособление к нередко встречающемуся в этих местообитаниях опылению с помощью летучих мышей (хироптерофилия): животным-опылителям - летучим мышам и летучим собакам - при подлете к дереву удобнее ухватываться за цветки.
Существенную роль в переносе пыльцы с цветка на цветок играют и птицы (это явление носит название "орнитофилия"). Орнитофильные растения заметны благодаря яркой окраске их цветков (красной, оранжевой, желтой), тогда как у хиропте рофильных растений цветки обычно невзрачные, зеленоватые или коричневатые.
Четкого различия между ярусами кустарников и трав, как это, например, характерно для лесов наших широт, во влажных тропических лесах практически не существует. Можно лишь отметить верхний ярус, в состав которого наряду с высокими крупнолистными представителями семейств банановых, марантовых, имбирных и ароидных входят кустарники и молодой подрост деревьев, а также нижний ярус, представленный низкорослыми, крайне теневыносливыми травами. По числу видов травянистые растения во влажном тропическом лесу уступают деревьям; но встречаются и такие равнинные влажные леса, не испытавшие воздействия человека, в которых вообще развит только один бедный видами ярус трав.
Обращает на себя внимание пока не нашедший объяснения факт пестролистности, а также наличия металлически-блестящих или матово-бархатистых участков поверхности у листьев растений, обитающих в припочвенном ярусе трав влажного тропического леса. Очевидно, эти явления в какой-то степени связаны с оптимальным использованием того минимума солнечного света, который доходит до таких местообитаний. Многие "пестролистные" растения нижнего яруса трав влажного тропического леса стали излюбленными комнатными декоративными растениями, например виды родов Zebrina, Tradescantia, Setcreasea, Maranta, Calathea, Coleus, Fittonia, Sanchezia, Begonia, Pilea и др. (рисунок на стр. 101). В глубокой тени доминируют разные папоротники, плаунки (Selaginella) и мхи; число их видов здесь особенно велико. Так, большинство видов плаунков (а их около 700) встречаются во влажных тропических лесах.
Напротив, гетеротрофные (то есть питающиеся готовыми органическими веществами) высшие растения встречаются здесь относительно редко. Правда, один из самых примечательных представителей растительного мира принадлежит именно к этой группе растений; речь идет о раффлезии (Rafflesia arnoldii) - растении с гигантскими, до метра в диаметре, цветками (рисунок на стр. 102). Это - растение-паразит, вегетативные части тела которого редуцированы до состоящих из клеток нитей, похожих на гифы (грибницу) грибов. Rafflesia arnoldii вместе с другими 11 видами рода Rafflesia, имеющими менее крупные цветки, встречается в Индо-Малайской области, где паразитирует на лианах из рода Cissus, относящегося к семейству Виноградовых.
Примечательны также живущие на почве влажных тропических лесов сапрофитные (то есть использующие разлагающиеся органические вещества) грибы семейств Clathraceae и Phallaceae. Они имеют своеобразные плодовые тела - "грибы-цветки" (см. рисунок на стр. 102).
Лианы. Если плыть через тропический влажный лес по реке, бросается в глаза обилие лиан (взбирающихся по деревьям растений с одревесневающими стеблями) - они, точно плотным занавесом, покрывают растущие по берегам деревья. Лианы - один из самых удивительных компонентов растительного покрова тропических районов: свыше 90% всех их видов встречается только в тропиках. Большинство растет во влажных лесах, хотя для своего развития они требуют хорошего освещения. Вот почему не везде они встречаются с одинаковой частотой. Прежде всего их можно видеть по лесным опушкам, на естественно образовавшихся осветленных участках леса и - по крайней мере иногда - в проницаемых для солнечных лучей ярусах древесных растений (см. рисунок на стр. 106). Особенно обильны они на плантациях, заложенных в области влажных тропических лесов, и во вторичных лесах, появляющихся на вырубках. В равнинных же влажных лесах, не испытавших на себе влияния человека, где густые, хорошо развившиеся кроны деревьев плотно сомкнуты, лианы встречаются сравнительно редко.
По способу закрепления на растениях, служащих им опорой, лианы можно подразделить на разные группы. Например, опирающиеся лианы могут удерживаться на других растениях с помощью опорных (цепляющихся) побегов или листьев, шипов, колючек или особых выростов типа крючков. Типичными примерами таких растений могут служить пальмы-ротанги рода Calamus, 340 видов которого распространены в тропиках Азии и Америки (см. рисунок на стр. 103).
Закрепляющиеся корнями лианы удерживаются на опоре с помощью множества мелких придаточных корешков или охватывают ее более длинными и толстыми корнями. Таковы многие теневыносливые лианы из семейства ароидных, например виды родов Philodendron, Monstera, Raphidophora, Syngonium, Pothos, Scindapsus, а также ваниль (Vanilla) - род из семейства орхидных.
Вьющиеся лианы охватывают опору сильно разрастающимися в длину междоузлиями. Обычно в результате последующего утолщения и одревеснения такие побеги закрепляются плотно. К группе вьющихся относится большинство тропических лиан, например представители богатых видами и распространенных по всем тропикам семейства мимозовых и родственного ему семейства цезальпиниевых, в частности энтада лазящая (Entada scandens); бобы последней достигают 2 м в длину (см. рисунок на стр. 104). К этой же группе принадлежат так называемая обезьянья лестница, или баугиния сассапарелевидная (Bauhinia smilacina), образующая толстые одревесневающие побеги, а также лианы с причудливыми цветками (виды кирказона, Aristolochia; семейство кирказоновых) (см. рисунок на стр. 103).
Наконец, прикрепляющиеся усиками лианы образуют одревесневающие усики - ими они цепляются за растения, служащие им опорой. К их числу относятся представители распространенного по всем тропикам рода Cissus из семейства Виноградовых, разные виды бобовых, в частности Physostigma venenosum (см. рисунок), а также виды страстоцвета (Passiflora; семейство страстоцветных).
Эпифиты. Чрезвычайно интересны приспособления к условиям существования во влажных тропических лесах у так называемых эпифитов - растений, живущих на деревьях. Число их видов очень велико. Они обильно покрывают стволы и ветви деревьев, благодаря чему оказываются достаточно хорошо освещенными. Развиваясь высоко на деревьях, они теряют возможность получать влагу из почвы, поэтому снабжение водой становится для них жизненно важным фактором. Не удивительно, что особенно много видов эпифитов там, где осадки обильны, а воздух влажен, но для оптимального их развития решающее значение имеет не абсолютное количество выпадающей влаги, а число дождливых и туманных дней. Неодинаковый микроклимат верхнего и нижнего древесного ярусов оказывается также причиной того, что обитающие там сообщества растений-эпифитов очень сильно различаются видовым составом. В наружных частях крон доминируют светолюбивые эпифиты, тогда как теневыносливые господствуют внутри, в постоянно влажных местообитаниях. Светолюбивые эпифиты хорошо приспособлены к смене сухих и влажных периодов времени, происходящей в течение суток. Как показывают приведенные далее примеры, для этого они используют разные возможности (рисунок на стр. 105).
У орхидных, представленных огромным числом видов (а большинство из 20 000-25 000 видов орхидей - это эпифиты), органами, запасающими воду и питательные вещества, служат утолщенные участки побегов (так называемые бульбы), листовые пластинки или корни. Такому образу жизни способствует также образование воздушных корней, которые снаружи покрыты слоями клеток, быстро поглощающих воду (velamen).
Растения влажного тропического леса, развивающиеся в припочвенном ярусе

Piled cadierei

Fittonia erschaffeltii
Episcia cupreata

Calathea ornata

Zebrina pendula
Семейство бромелиевых, или ананасовых (Bromeliaceae), представители которого распространены, за одним исключением, в Северной и Южной Америке, состоит почти только из эпифитов, чьи розетки листьев, похожие на воронки, служат водосборными резервуарами; из них вода и растворенные в ней питательные вещества могут всасываться чешуйками, находящимися у оснований листьев. Корни же служат только как органы, прикрепляющие растения.
Даже кактусы (например, виды родов Epiphyllum, Rhipsalis, Hylocereus и Deamia) в горных влажных тропических лесах растут как эпифиты. За исключением немногих видов рода Rhipsalis, встречающихся также в Африке, на Мадагаскаре и в Шри Ланке, все они растут только в Америке.
Некоторые папоротники, например, папоротник- птичье гнездо, или асплениум гнездовой (Aspleniumnidus), и папоротник-оленьи рога, или платицериум оленерогий (Platycerium), благодаря тому что у первого листья образуют воронкообразную розетку, а у второго имеются специальные листья, прилегающие к стволу дерева-опоры, подобно накладным карманам (рисунок на стр. 105), даже способны создавать похожий на почву, постоянно влажный субстрат, в который врастают их корни.
Эпифиты, развивающиеся в затененных местообитаниях, представлены прежде всего так называемыми гигроморфными папоротниками и мхами, которые приспособились к существованию во влажной атмосфере. Наиболее характерные компоненты таких сообществ эпифитных растений, особенно ярко выраженных в горных влажных лесах, - это гименофилловые, или тонколистниковые, папоротники (Hymenophyllaceae), например, представители родов Hymenophyllum и Trichomanes. Что же касается лишайников, то они из-за медленного роста не играют столь большой роли. Из цветковых растений в этих сообществах встречаются виды родов Peperomia и Begonia.
Даже листья, и прежде всего листья деревьев нижних ярусов влажного тропического леса, где постоянно высока влажность воздуха, могут быть заселены разными низшими растениями. Это явление называют эпифиллией. Поселяются на листьях преимущественно лишайники, печеночные мхи и водоросли, образующие характерные сообщества.
Своеобразную промежуточную ступень между эпифитами и лианами представляют собой гемиэпифиты. Они либо растут сначала как эпифиты на ветвях деревьев, а по мере образования воздушных корней, доходящих до почвы, становятся самостоятельно укрепляющимися в почве растениями, либо на ранних стадиях развиваются как лианы, но затем теряют связь с почвой и таким образом превращаются в эпифиты. К первой группе относятся так называемые деревья-душители; их воздушные корни, словно сетью, охватывают ствол дерева-опоры и, разрастаясь, настолько препятствуют его утолщению, что дерево в конце концов отмирает А совокупность воздушных корней становится после этого как бы системой "стволов" самостоятельного дерева, на ранних стадиях развития бывшего эпифитом. Наиболее характерными примерами деревьев-душителей могут служить в Азии виды рода Ficus (семейство тутовых), а в Америке - представители рода Clusia (семейство зверобойных). Ко второй группе относятся виды семейства ароидных.
Паразиты и сапрофиты влажного тропического леса
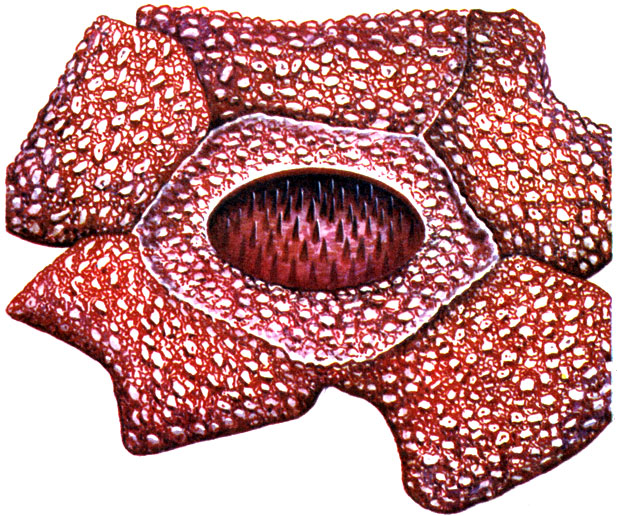
Rafflesia arnoldii

Clathrus chrysomycelinus
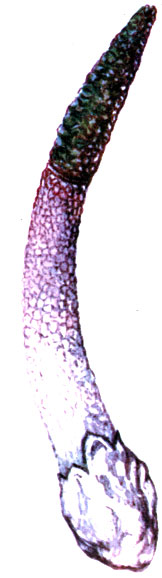
Mutinus bambusinus
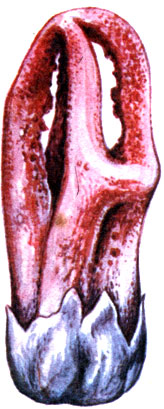
Laternea columnata
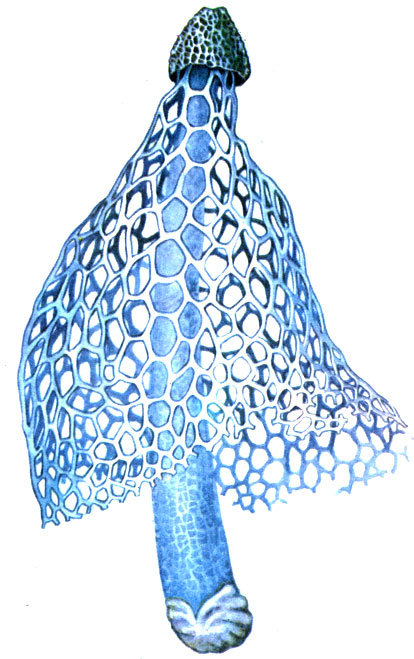
Schleierdame Dictyophora indusiata
Вечнозеленые влажные тропические леса низменностей. Хотя флористический состав влажных тропических лесов в разных районах земного шара весьма различен, и три основные области таких лесов обнаруживают в этом отношении лишь незначительное сходство, все же в характере их растительного покрова повсюду можно выявить аналогичные модификации основного типа.
Прототипом влажного тропического леса считают вечнозеленый влажный тропический лес незатопляемых низменностей, не бывающих продолжительное время сырыми. Это, так сказать, нормальный тип леса, о структуре и особенностях которого мы уже говорили. Лесные сообщества речных пойм и затопляемых низин, а также болот отличаются от него обычно менее богатым видовым составом и присутствием растений, которые приспособились к существованию в таких местообитаниях.
Пойменные влажные тропические леса встречаются в непосредственной близости к рекам на регулярно затопляемых территориях. Они развиваются, в местообитаниях, образовавшихся в результате ежегодного отложения богатых питательными веществами речных наносов - принесенных рекой взвешенных в воде и затем осевших мельчайших частиц. Эту мутную воду так называемые "беловодные" реки приносят преимущественно из безлесных районов своих бассейнов*. Оптимальное содержание питательных веществ в почве и относительная обеспеченность проточной воды кислородом обусловливают высокую продуктивность растительных сообществ, развивающихся в таких местообитаниях. Пойменные тропические леса труднодоступны для освоения их человеком, поэтому они и поныне в основном сохранили свою первозданность.
* (Реки, авторами этой книги называемые "беловодными", в Бразилии принято называть белыми (rios blancos), а "черновод- ными" - черными (rios negros). Белые реки несут мутную воду, богатую взвешенными частицами, но цвет воды в них может быть не только белым, но и серым, желтым и т. д. Вообще для рек бассейна Амазонки характерно удивительное разнообразие окраски вод. Черные реки обычно глубокие; в них воды прозрачные - они кажутся темными только потому, что в них нет взвешенных частиц, отражающих свет. Растворенные в воде гумусовые вещества лишь усиливают этот эффект и, по-видимому, влияют на оттенок окраски.)

Лианы на опушке горного дождевого тропического леса
Лианы влажного тропического леса
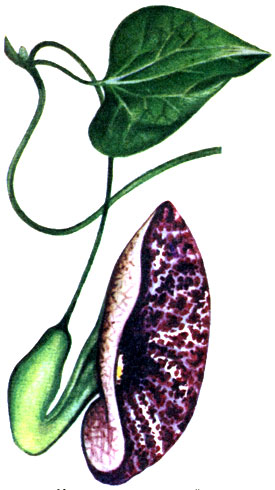
Кирказон изящный Aristolochia elegans

Physostigma venenosum

Монстера Monstera deliciosa

ВанильVanilla planifolia

Пальма-ротанг Calamus niger

Спирально закрученные стебли энтады лазящей (Entada scandens) - лианы влажного тропического леса
Продвигаясь от самого берега реки поперек поймы до ее края, можно выявить характерную последовательность растительных сообществ, обусловленную постепенным понижением уровня поверхности почвы от высоких прирусловых валов к краю поймы. На редко затапливаемых прирусловых валах растет богатый лианами прирусловый лес, далее от реки переходящий в настоящий заливаемый лес. У дальнего от берега края поймы встречаются озера, окруженные тростниковыми или травяными болотами.

Проточное пойменное озеро с сообществами водных растений и затопляемый лес в бассейне Амазонки. На переднем плане - плодоносящее калебасовое дерево (Crescentia cujete)
Болотистый дождевой лес. В местообитаниях, почвы которых почти постоянно покрыты стоячей или медленно текущей водой, растут болотистые тропические дождевые леса. Их можно обнаружить преимущественно около так называемых "черноводных" рек, истоки которых находятся в облесенных районах. Поэтому их воды не несут взвешенных частиц и имеют окраску от оливковой до черно-коричневой из-за содержания в них гумусовых веществ. Самая известная "черноводная" река - Риу Негру, один из важнейших притоков Амазонки; она собирает воду с огромной территории, имеющей подзолистые почвы.
В отличие от пойменного влажного тропического леса болотистый лес, как правило, покрывает всю долину реки. Здесь происходит не отложение насосов, а, напротив, лишь равномерное вымывание, поэтому поверхность долины такой реки ровная.
Из-за необеспеченности местообитаний питательными веществами болотистые дождевые леса не столь пышны, как пойменные, а из-за недостатка воздуха в почве здесь часто встречаются растения с воздушными и ходульными корнями. По этой же причине разложение органических веществ происходит медленно, что способствует образованию мощных торфоподобных слоев, чаще всего состоящих из более или менее разложившейся древесины.
Полувечнозеленые влажные леса низменностей. Для некоторых областей дождевых тропических лесов характерны короткие засушливые периоды, вызывающие смену листвы у деревьев верхнего яруса леса. При этом нижние древесные ярусы остаются вечнозелеными. Такая переходная ступень к облиствленным в период дождей сухим лесам (см. стр. 120) получила название "полувечнозеленые, или полулистопадные, влажные леса низменностей". В засушливые периоды здесь может происходить передвижение влаги в почве снизу вверх, поэтому такие леса получают достаточно питательных веществ и оказываются весьма продуктивными.
Эпифиты тропического влажного леса

Rhipsalis cassutha

Платицериум оленерогий Platycerium alcicorne

Сверху Асплениум гнездовой Asplenium nidus и снизу Cattleya citrina

Vriesea splendens
Горные тропические влажные леса. Описанным выше лесам, существование которых определяется присутствием воды, можно противопоставить такие варианты влажного тропического леса, образование которых связано с понижением температуры; они в основном встречаются на влажных местообитаниях, находящихся в разных высотных поясах горных областей тропических регионов. В предгорном поясе, на высоте примерно 400-1000 м над уровнем моря, влажный тропический лес почти не отличается от леса низменностей. В нем имеются только два яруса деревьев, а деревья верхнего яруса не такие высокие.

Горный тропический влажный лес на острове Гренада (Малые Антильские острова)
Зато тропический влажный лес горного пояса, или, как принято говорить, горный влажный лес, растущий на высоте 1000-2500 м, обнаруживает более существенные отличия. Он также имеет два древесных яруса, но часто их трудно выявить, а верхняя их граница нередко не превышает 20 м. Кроме того, здесь меньше видов деревьев, чем во влажных лесах низменностей, отсутствуют и некоторые характерные особенности деревьев таких лесов, в частности и ходульные корни, а также каулифлория. Листья деревьев обычно более мелкие и не имеют заострений для удаления капель воды.
В ярусах кустарников и трав часто преобладают папоротники и виды бамбука. Очень обильны эпифиты, тогда как крупные лианы встречаются редко.
На еще больших высотах постоянно влажных тропиков (2500-4000 м) горные влажные леса сменяются субальпийскими горными лесами, развивающимися на уровне облаков (см. т. 2).
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'