Спорообразование у анаэробных бактерий
Споры представляют собой специфическим образом устроенные покоящиеся зародышевые клетки, выдерживающие влияние высокой температуры, радиации, вакуума, различного рода токсических веществ и других неблагоприятных факторов, приводящих к гибели вегетативные клетки.
Секрет их необычной устойчивости давно интригует ученых, и в последние годы во многих странах развернулись интенсивные исследования процесса спорообразования.
Бактерргальные споры образуются эндогенно, т. е. внутри материнских вегетативных клеток. Что заставляет бактерии перейти от вегетативного размножения к спорообразованию? Формирование спор наступает на определенной стадии развития в тот момент, когда в среде исчерпываются пищевые ресурсы (прежде всего источники углерода и азота) и происходит накопление токсичных продуктов обмена веществ. Каков же конкретный пусковой механизм, включающий процесс спорообразования? На этот вопрос ответа пока нет.
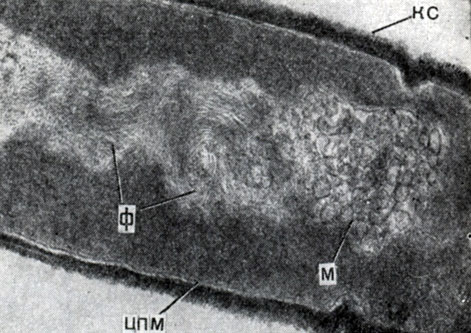
Рис. 43. Ультратонкий срез клеток Clostridium penicillum. Видны клеточная стенка (КС), нуклеопд со спирально закрученными фибриллами ДНК(Ф), связанные с нуклеоидом мезосомы (М), и цитоплазматическая мембрана (ЦПМ)
Основное назначение спорообразования - перевести культуру в покоящееся (анабиотическое) состояние. Действительно, у зрелых спор обмен веществ находится на крайне низком уровне. Это позволяет бактериям сохраниться при неблагоприятных условиях среды, а при изменении условий в благоприятную сторону снова перейти к вегетативному росту. Для анаэробов (особенно почвенных) крайне важно также, что споры не чувствительны к кислороду. Это позволяет им выжить в аэробных условиях, губительно действующих на вегетативные клетки. Способность бактерий образовывать споры позволяет им оставаться жизнеспособными в течение десятков и сотен лет. Жизнеспособные споры были выделены из египетских мумий, трупов мамонтов и других объектов, где они провели века и тысячелетия.
Таким образом, споры служат целям сохранения вида.
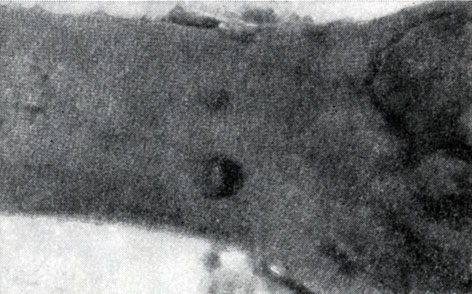
Рис. 44. Ультраструктура клеточной стенки у Clostridium sp. Увел. X 45 000
Цитология спорообразования. Молодые интенсивно делящиеся клетки анаэробов содержат нуклеоиды в виде гантелек, или V-образных фигур (рис. 46). Перед спорообразованием деление клеток прекращается, они резко увеличиваются в размерах. В это время происходит накопление большого количества запасного питательного вещества - гранулезы, - откладывающегося в виде гранул, из-за чего цитоплазма становится зернистой, а сами клетки раздуваются, принимая вид лимона (клостридии) (рис. 47 - 49) либо барабанной палочки (плектридии) (рис. 50, 51). Лишь у части про-теолитических анаэробов клетки не меняют своего первоначального вида, сохраняя обычную палочковидную (бациллярную) форму (рис. 52).
Первым признаком наступления спорообразования является изменение морфологии нуклеоидов, принимающих вид шаровидных телец. Далее несколько нуклеоидов сближаются на одном из полюсов клетки, сливаются и образуют продольно расположенный извитый хроматиновый (ядерный) тяж (рис. 53, 54 и схема 1 на табл. 32).
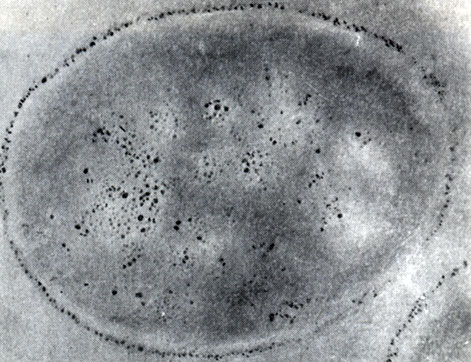
Рис. 45. Включения гранулезы в клетках Clostridium taeniosporum. 'Окраска' полисахаридов серебром. От ложения зерен серебра видны в светлых шаровидных включениях и в клеточной стенке. Увел. X 45 000
Зона цитоплазмы, в которой расположен ядерный тяж, превращается далее в проспору. У бактерий с мелкими клетками перед спорообразованием имеются обычно 2 отдельных нуклеоида, которые сливаются с образованием осевой хроматиновой нити. Впоследствии только часть этой нити переходит в спору. Третий тип поведения ядра встречается у многих сахаролитических анаэробов. Ядерное вещество у них имеет вид хроматиновой сеточки, расположенной по всей цитоплазме. Часть этой сеточки стягивается на одном из полюсов клетки с образованием тяжа - центра формирующейся проспоры. С помощью обычного микроскопа можно выделить три стадии формирования споры. Первая - возникновение на одном из полюсов клетки спорогенной зоны, в которой хорошо заметно ядерное вещество в виде светлых палочек (рис. 39). Вторая - спорогенная зона превращается в темную (оптически плотную) овальной формы проспору с четко выраженными контурами. В проспорах ядерное вещество уже не выявляется без применения специальных методов (окрашивание). Третья - проспоры постепенно светлеют, приобретая способность сильно преломлять свет, и теряют способность окрашиваться красителями. Созревшие споры выглядят светлыми, резко преломляющими свет тельцами, обладающими мощной оболочкой (рис. 40). Форма зрелых спор может быть различной у разных видов анаэробов: сферическая, овальная, яйцевидная, цилиндрическая.
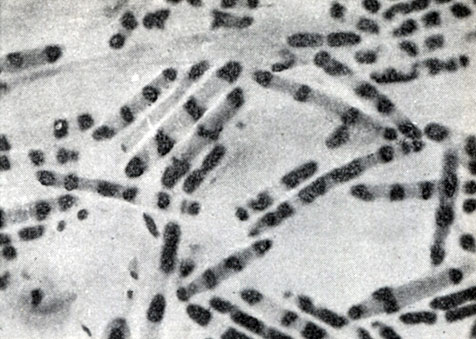
Рис. 46. Молодые, интенсивно делящиеся клетки Clostridium sporopenitum. Видны делящиеся нуклеоиды. Окраска ядерного вещества по методу Романовского - Гимза. Увел. X 3500
В люминесцентном микроскопе при окраске акридиновым оранжевым (люминесцирующий краситель) наблюдается многокрасочное изображение. Вегетативные клетки светятся (люминесцируют) слабым зеленоватым светом. Их ядра ярко люминесцируют зеленым светом (табл. 32, схема 1). Ядерные тяжи в спорогенной зоне сначала зеленые, затем окружаются веществом с красной флюоресценцией. Далее флюоресцирующая красная зона принимает овальную форму, а хроматиновый, расположенный в центре тяж светится желтым светом. При созревании спора резко меняет цвет и начинает светиться ярким зеленым светом. У зрелых спор с хорошо выраженным кортексом (корой) сердцевина споры темная, зеленым светится только оболочка. Меняется также и цвет цитоплазмы материнской клетки (спорангия). Сначала она слабо-зеленого цвета, затем, с момента оформления проспоры, начинает светиться оранжевым светом.
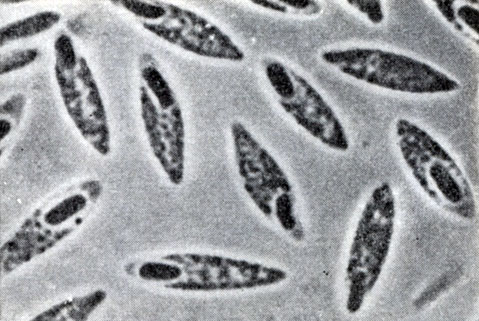
Рис. 47. Клострпдиальная форма клеток Clostridium butyricum. Увел. X 3500
Хроматиновое вещество сначала выглядит в виде компактных шаровидных гранул, а при созревании проспоры заметно разрушение нуклеоидов.
Биологическое значение слияния ядерного вещества перед спорообразованием остается неясным. Слияние хроматина наблюдается и у неспорулирующих вегетативных клеток под влиянием антибиотиков, низкой температуры, высоких концентраций солей и т. д. Во всех этих случаях, как и при спорообразовании, происходит остановка или резкое снижение синтеза ДНК. Отсюда некоторые исследователи делают вывод об отсутствии специфического значения слияния ядерного вещества перед спорообразованием. Другие авторы считают, что подобное поведение ядра имеет определенный генетический смысл. Интересно, что не только цитологические, но и химические анализы также свидетельствуют о необычном поведении ДНК при спорообразовании. Например, в спорах анаэробов нуклеотидный состав ДНК смещается по сравнению с ДНК вегетативных клеток в сторону ГЦ-типа (ДНК в спорах более "богата" такими основаниями, как гуанин и цитозин). Для спорообразующих аэробов, кроме того, показано, что около 40% высокополимерной ДНК перед спорообразованием выделяется из клеток в среду (редукция части хроматина).
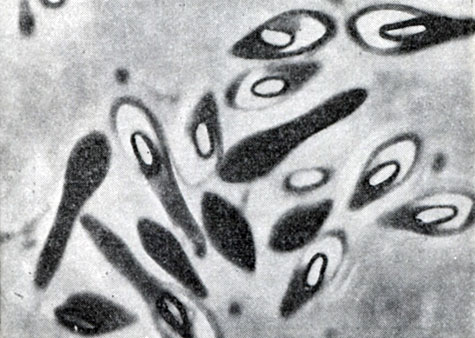
Рис. 48. Клострпдиальная форма клеток. Споры снабжены светлыми конусовидными колпачками. Clostridium species, шт. 1. Увел. X 3500
Электронная микроскопия с ее высокой разрешающей способностью открывает много новых деталей процесса спорообразования. Нужно иметь в виду, что принципы формирования изображения в электронном и обычном световом микроскопе значительно отличаются: в обычном используется видимый свет (фотоны), а в электронном - поток отрицательно заряженных частиц - электронов.
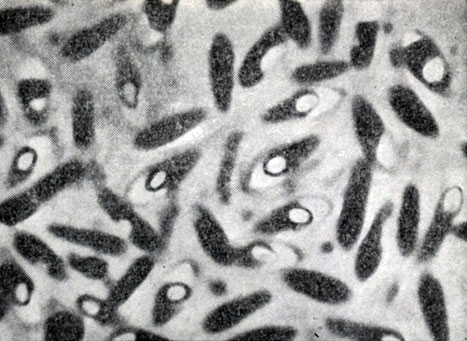
Рис. 49. Клостридиальная форма клеток при спорообразовании. Споры со светлыми конусовидными колпачками. Clostridium sp. Увел. X 3500
Эти различия необходимо учитывать, чтобы правильно оценивать электронно-микроскопические снимки. В обычном микроскопе изображение создается главным образом за счет различий в степени поглощения света разными участками исследуемого объекта, в электронном - в основном за счет рассеяния объектом электронов. Проходя через объект, электроны сталкиваются с атомными ядрами, рассеиваются и задерживаются. Только та часть электронов, которая беспрепятственно пройдет через объект и попадет на флюоресцирующую пластинку (экран микроскопа), сможет создать изображение. Те места на экране, куда попадут электроны, будут светиться. То есть участки клеток, сильно рассеивающие электроны, будут выглядеть на экране темными, а участки, слабо рассеивающие электроны, - светлыми.
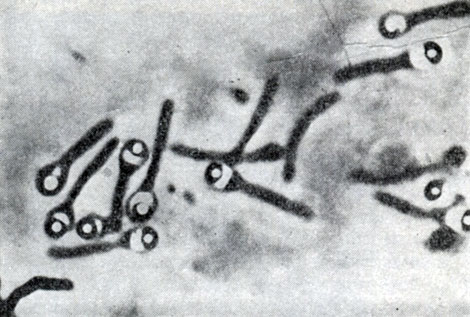
Рис. 50. Плектридиальная форма клеток при спорообразовании. Споры с серповидными колпачками. Clostridium sp. Увел. X 3500
Биологические объекты (клетки) состоят из веществ, построенных главным образом из легких элементов (С, N, О, Н, Р, S и др.), поэтому их изображение в электронном микроскопе слабо контрастно - в клетках можно увидеть очень мало структурных деталей. При использовании светового микроскопа это затруднение преодолевается при помощи окрашивания (контрастирования) объектов различными красителями. В электронном микроскопе изображение одноцветное. Чтобы сделать изображение более контрастным, клетки обрабатывают солями тяжелых металлов (свинца, ртути, хрома, урана, вольфрама). Так как атомы тяжелых металлов очень сильно рассеивают электроны, то структуры клетки, поглотившие эти металлы, будут выглядеть темными и контрастными. Соли тяжелых металлов или их окислы принято теперь называть электронными красителями.
Рис. 51. Плектридии, адсорбированные на целлюлозном волокне. (Клетчаткоразрушающне бактерии.) Увел. X 3500
Электронную микроскопию в настоящее время нельзя рассматривать только как более утонченный морфологический метод, так как при описании электронно-микроскопических изображений цитологи неизбежно переходят с описания формы на язык химии и физической химии и пытаются объяснить на молекулярном уровне строение и функцию тех или иных структур клетки. Поэтому морфология, "спустившись на субмикроскопический уровень, органически срастается с биохимией и физиологией клетки" (Г. М. Франк).
Рис. 52. Бациллярный тип спорообразования у Clostridium sporopenatum. Увел. X 3500
Целые клетки из-за своей большой толщины непрозрачны для пучка электронов. Поэтому, чтобы выявить внутреннюю структуру, бактериальную клетку разрезают на десятки и даже сотни отдельных ломтиков (срезов) толщиной всего 200 - 600 А. Далее эти срезы просматривают в микроскопе.
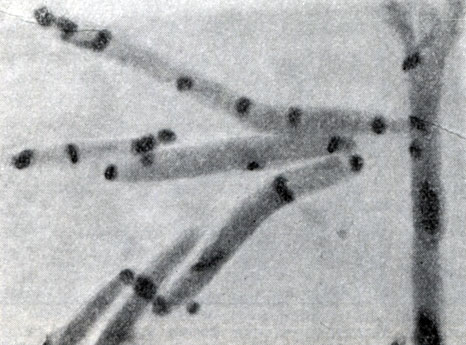
Рис. 53. Слияние нуклеоидов на полюсах клеток (перед спорообразованием). Clostridium sporopenitum Увел. X 3500
Электронно-микроскопические исследования ультратонких срезов спорулирующих клеток бактерий показали, что формирование проспоры начинается с инвагинации (врастания) цитоплазматической мембраны ближе к одному из полюсов клетки (схема 2 на табл. 32). При этом мембрана продвигается к центру клетки, и полюса ее сливаются с образованием споровой перегородки (септы). В этом процессе участвуют мезосомы, которые как бы спаивают сближающиеся участки инвагинированных мембран (рис. 1 на табл. 33). Септа состоит из двух элементарных мембран. На этом заканчивается вторая стадия спорообразования (если за первую принять образование хроматинового тяжа). Эту стадию можно рассматривать как модифицированное клеточное деление, которое, как известно, также происходит благодаря инвагинации цитоплазматической мембраны и образования септы.
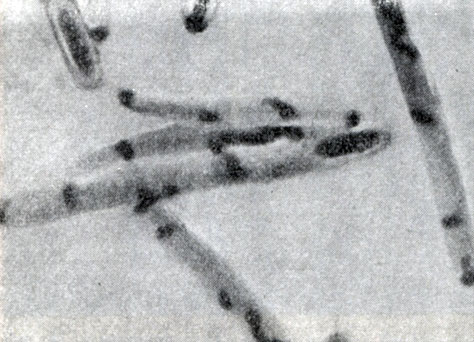
Рис. 54. Образование ядерных тяжей в процессе спорообразования. Clostridium sporopenitum. Увел. X 3500
Следующей стадией является процесс "поглощения" материнской клеткой септированного (отсеченного) участка цитоплазмы с ядром. Этот процесс осуществляется путем роста и продвижения периферических участков мембраны материнской клетки по направлению к полюсу клетки. Затем сближающиеся участки мембраны сливаются и образуется проспора, обладающая двумя элементарными (трехслойными) мембранами - внутренней и внешней (рис. 2 на табл. 33). У одних видов проспора остается в дальнейшем у полюса клетки (терминальное расположение), у других она перемещается внутрь цитоплазмы, занимая центральное либо субтерминальное положение. Таким образом, в конце этой стадии образуется своеобразный двуклеточный организм: внутри цитоплазмы материнской клетки возникает новая клетка - проспора, окруженная, в отличие от материнской, двумя элементарными мембранами. С этого момента начинается новая необратимая фаза развития и метаболизма, заканчивающаяся созреванием споры и гибелью материнской клетки.
Интересно, что, в отличие от четвертой, вторая и частично третья стадии спорообразования обладают еще свойством обратимости. Так, если после образования септы добавить к спорулирующей культуре антибиотик хлорамфеникол, то синтез белков (а значит, и рост мембраны) будет подавлен. Движение периферических участков мембраны, поглощающей отсеченный участок протопласта, будет остановлено. В результате начавшийся процесс спорообразования превратится в обычный процесс вегетативного деления клеток, и между двумя мембранами септы отложится материал клеточной стенки (рис. 55), чего не бывает при нормальном течении спорообразования. Считается, что на четвертой стадии споруляции происходит образование кортикального слоя (кортекса) между внутренней и внешней мембранами проспоры. Сначала кортекс появляется в виде тонкого темного слоя, сходного по структуре и плотности с клеточной стенкой вегетативной клетки. Затем этот слой резко увеличивается в толщине за счет образования более электронно-прозрачного (светлого) слоя (рис. 2 - 5 на табл. 33).
На пятой стадии закладывается оболочка споры. Вначале вокруг проспоры на некотором расстоянии от внешней мембраны проспоры в цитоплазме материнской клетки возникают участки темного (электронно-плотного) вещества в виде чешуек. На шестой стадии отдельные листки оболочки удлиняются и, в конце концов, сливаются, образуя сплошной непрерывный плотный слой. Между этим слоем и внешней мембраной проспоры остается отсеченный слой цитоплазмы вегетативной клетки. Поверх первого слоя оболочки может закладываться еще один или два слоя. В этом случае они разделяются на внутренний, средний и внешний слои оболочки. Эти слои различаются между собой по структуре. У некоторых видов внутренний слой оболочки является пластинчатым, а внешний имеет вид плотного толстого слоя. У других видов, наоборот, пластинчатый слой может быть внешним, а более плотный слой - внутренним. Если строение сердцевины очень сходно у разных видов, то строение оболочек спор у них сильно различается как по тонкому строению, так и по количеству и толщине слоев (рис. 56).
После окончательного созревания споры происходит лизис материнской вегетативной клетки - клеточная стенка разрушается, и спора выходит в среду (седьмая стадия).
У многих видов анаэробов поверх споровой оболочки обнаруживается еще одна структура - экзоспориум. Он имеет вид чехла, в котором расположена спора (рис. 57). Тонкое строение экзоспориума весьма различно у разных видов (рис. 56). Часто он очень многослоен, например у Clostridium pasteurianum, CI. bifermentans, CI. tyrobutyricum и др. В экзоспориуме многих видов анаэробов обнаруживаются слои с упорядоченно расположенными субъединицами. На рисунке 58 показана тонкая структура пластинчатого слоя экзоспориума. Шаровидные субъединицы в этом слое имеют гексагональную упаковку. Иногда рядом расположенные субъединицы сливаются, образуя кольцевидные структуры с порами в центре. Такие слои экзоспориума представляют собой перфорированные пленки-мембраны. Возникает экзоспориум на ранней стадии формирования споры в виде небольшого пузырька на внешней мембране проспоры. Этот пузырек разрастается, превращаясь в чехол, охватывающий спору со всех сторон.
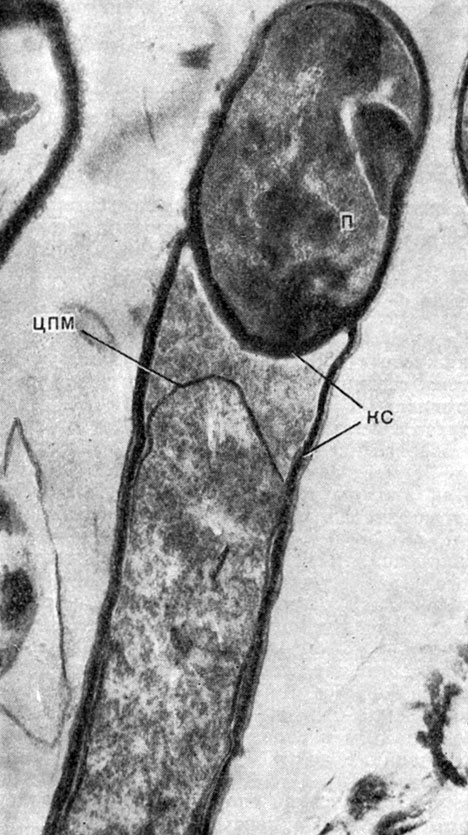
Рис. 55. Влияние хлорамфеникола (антибиотика, подавляющего синтез белка) на спорообразование. Остановка на стадии образования септы и превращение спорообразования в процесс вегетативного деления клеток. Увел. X 66 000. Clostridium penicillum. П - проспора, ЦПМ - цитоплазматическая мембрана, КС - клеточная стенка. Увел, Х 66 000
У анаэробов этот чехол не цельный, имеет крупные поры. Иногда через эти поры в цитоплазму спорангия проходят трубчатые выросты споровой оболочки.
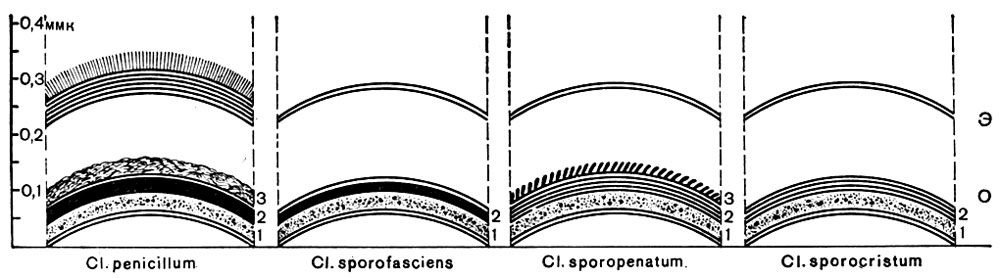
Рис. 56. (Схема.) Строение оболочки спор у различных видов анаэробных бактерий: Э - экзоспориум, О-оболочка споры. 1, 2, 3 - слои споровой оболочки
Химический состав и роль споровых структур. Сердцевина споры, окруженная слоем кортекса, представляет собой протопласт с собственной мембраной, ядром и цитоплазмой (схема 3 на табл. 32). Сердцевина у зрелых спор является покоящейся вегетативной клеткой. Она характеризуется очень низким уровнем метаболизма. Хотя в ней имеются все необходимые ферменты, их активность каким-то образом подавлена.
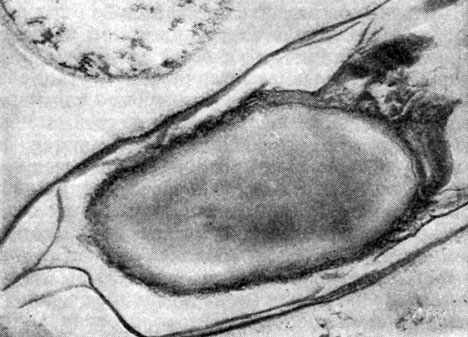
Рис. 57. Зрелая спора Clostridium penicillum. Сердцевина выглядит бесструктурной. Видны многослойный экзоспориум и выросты на оболочке споры. Увел. X 45 000.
Кортекс (схема 3 на табл. 32) состоит из мукопептидов, весьма сходных с мукопептидами клеточных стенок. В кортексе содержится также диаминопимелиновая кислота. Внутренняя плотная часть кортекса, прилегающая к мембране сердцевины, при прорастании спор оформляется в клеточную стенку молодой вегетативной клетки. В спорах обнаруживается в довольно больших количествах дипиколиновая кислота (C7H504N) - активный хелирующий агент, соединение, образующее клешневидные комплексы с металлами. Это вещество отсутствует в вегетативных клетках. Из спор дипиколиновая кислота выделяется в виде кальциевой и магниевой солей, играющих большую роль в термоустойчивости спор. Хорошо известно, что споры способны выдерживать высокие температуры и не погибают даже при кипячении. Имеются также доказательства того, что дипиколиновая кислота играет центральную роль в процессе перевода протопласта споры в покоящееся состояние. Но механизм этих явлений не выяснен. Предполагается, что дипиколиновая кислота локализована в кортексе, так как имеется определенная корреляция между образованием кортекса и накоплением в споре дипиколиновой кислоты и кальция. С другой стороны, при разрушении кортекса (в процессе прорастания или механическом разрушении спор) дипиколиновая кислота обязательно переходит в раствор.

Рис. 58. Тонкая структура экзоспориума у Clostridium bifermentans. Видны отдельные светлые субъедишщы и гексагональные 'плитки'. Увел. X 384 000
Кортекс у зрелых спор, очевидно, играет защитную роль. Он предохраняет сердцевину от литических ферментов, разрушающих клетки. Это предположение подтверждается экспериментами с мутантами, у которых потеряна способность к формированию кортекса. На завершающей стадии спорообразования происходит резкое возрастание активности литических ферментов, полностью разрушающих материнскую вегетативную клетку. Лишенные кортекса споры при этом также лизируются.
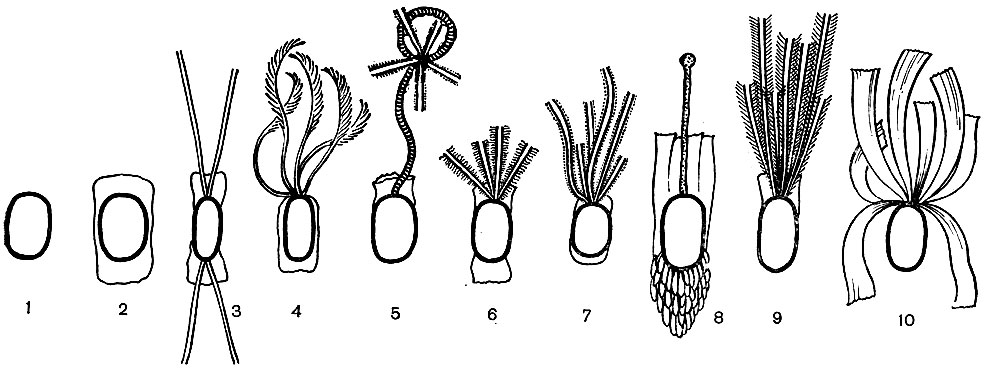
Рис. 59. Строение поверхностных структур спор некоторых анаэробных бактерий: Споры без выростов и экзоспориума: 1 - Clostridium felsineum. Споры с экзоспориумом в, виде чехла: 2 - Gl. pasteurianum. Споры с выростами различных типов и экзоспориумом: з - Gl. sporosetosum; 4 - CI. sporopenatum; 5 - Gl. saprogenes; 6 - CI. sporofasciens; 7 - Gl. sartagoformum; 8 - Gl. corinoforum; 9 - Gl. penicillum. Споры с лентовидными выростами, но без экзоспориума: 10 - Gl. taeniosporum
Оболочка (или покровы) является уникальной структурой бактериальных спор, не встречающейся у других микроорганизмов (схема 3 на табл. 32). Она в основном состоит из белковых веществ, обогащенных цистином. Объем оболочки достигает 50% от всего объема споры. Вещество споровых оболочек не чувствительно к действию различных литических ферментов. Специфическая для каждого вида бактерий форма спор сохраняется благодаря структурной ригидности оболочек. Оболочка играет также роль защитной структуры, предохраняющей споры от преждевременного прорастания. Споры мутантных штаммов, лишенные оболочек, обычно прорастают непосредственно после выхода из спорангиев в неблагоприятной для роста среде (даже в дистиллированной воде), что приводит к гибели проросшие клетки. Однако роль оболочек спор, так же как и кортекса, остается во многом еще загадочной.
Экзоспориум представляет собой мембрановидную, часто многослойную структуру. Химический состав экзоспориума проанализирован лишь у одной аэробной культуры. О составе экзоспориума у анаэробов данных в литературе нет. Предполагается, что экзоспориум играет роль барьера, регулирующего проникновение различных веществ в спору. В этом отношении необъясненным остается тот факт, что у многих анаэробных бактерий экзоспориум не представляет собой замкнутой системы: в его полярной части, погруженной в цитоплазму материнской клетки, имеются очень крупные поры, или отверстия, диаметром до 0,5 мкм. Некоторые авторы предполагают, что экзоспориум участвует в формировании споровых оболочек. После механического удаления экзоспориума споры остаются нормальными, процесс прорастания у них не нарушается. Роль экзоспориума продолжает оставаться загадкой.
Выросты на спорах. Замечательной особенностью спорообразования у анаэробов является формирование на спорах особых выростов с поразительно разнообразной структурой. Каждому виду анаэробных бактерий свойствен свой тип строения выростов (рис. 59). Этот признак строго специфичен, наследственно закреплен и очень устойчив. Даже у дефектных спор, потерявших способность образовывать оболочку, выросты сохраняются и не меняют своей специфической структуры.
Поражает разнообразие выростов у различных видов анаэробных бактерий. Они могут иметь форму метелок, тонких нитей или жгутиков, трубок различной толщины, разбросанных по всей поверхности споры или собранных в пучок ершиковидных палочек, длинных широких лент, шипов, булавок и образований, сходных по форме с оленьими рогами (табл. 34).
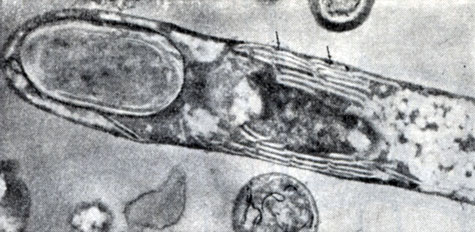
Рис. 60. Ультратонкий срез клеток CI. taeniosporum. Реакция на дегидрогеназы. Стрелками показаны отложения теллура. Видны выросты и светлые шаровидные включения гранулезы. Увел. X 45 000
У Clostridium taeniosporum выросты имеют лентовидную форму. Пучок таких отростков прикрепляется к споре с помощью специального органа - подушечки (схема 3 на табл. 32 и рис. 4 на табл. 35). Выросты появляются на ранней стадии формирования проспоры - перед закладкой кортекса и оболочки; затем они растут, удлиняются, пронизывая цитоплазму, пока не достигнут противоположного полюса клетки. Цитоплазма вокруг выростов постепенно лизируется (рис. 61). Материнская клетка разрушается. На свободных зрелых спорах, вышедших из спорангия, выросты распускаются в виде зонтика (рис. 3 на табл. 34).
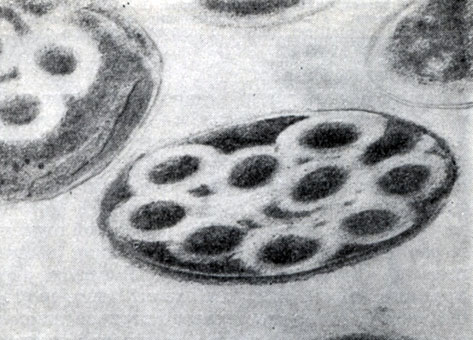
Рис. 61. Поперечный ультратонкпй срез клеток С1. penicillum. Хорошо заметен лизис цитоплазмы вокруг трубчатых выростов (светлые зоны). Увел. X 96 000
На спорах Clostridium saprogenes имеется по одному крупному и сложноустроенному выросту (табл. 35). Он имеет вид длинного толстого жгута-ствола, образующего на конце кольцо, от которого отходят усики - трубчатые палочковидные отростки. Ствол имеет грубозернистую структуру и поперечную исчер-ченность, мелкозернистые усики обладают капсул ярным слоем. Формирование выростов у этого вида можно проследить на целых клетках. Сначала отростки плохо просматриваются, так как они окружены плотными участками цитоплазмы (рис. 1 на табл. 35), затем цитоплазма осветляется и выросты становятся хорошо заметными (рис. 2 на табл. 35). На одном из полюсов клетки четко заметна кольцевидная структура и усики.
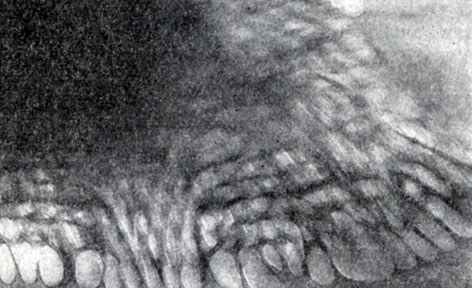
Рис. 62. Ячеистые колпачки на спорах. Видны отдельные газовые вакуоли (мешочки), составляющие колпачок. G1. caliptrosporum. Увел. X 55 000
Следующая стадия - лизис материнской клетки и выход зрелой споры в среду (рис. 3 на табл. 35). У организма, описанного как Bacillus (Clostridium) penicillus (строгий анаэроб), выросты имеют вид пучка толстых трубок, прикрепленных к одному из полюсов споры (рис. 1 на табл. 34). На поперечных срезах спорулирующих клеток четко видны зоны лизиса цитоплазмы вокруг выростов, имеющих вид овальных плотных телец с зернистой структурой (рис. 61).
Химической состав и роль выростов на спорах. Сложные по строению отростки на спорах впервые были обнаружены сотрудниками кафедры биологии почв Московского университета. В настоящее время выросты на спорах анаэробов изучаются также во многих лабораториях за рубежом. Описываются новые виды отростков. Наиболее интересным является вопрос о функции (биологической роли) этих споровых структур. Выяснение функции тех или иных клеточных структур - отнюдь не легкое дело. В истории биологии найдется немало примеров тому, что выяснение роли той или иной клеточной структуры занимает много лет (а иногда и десятилетий). Подобные исследования очень увлекательны и имеют большое значение: с каждой новой структурой открываются новые вещества, особые ферменты, нередко совершенно неизвестные биохимические процессы.
Функция выростов на спорах окончательно еще не выяснена. Одни исследователи предполагают, что отростки на спорах - специфические чувствительные (хемосенсорные) органел-лы, подающие споре "команду" на прорастание (при благоприятных условиях). Другие считают, что выросты выполняют важную роль в процессе созревания спор, участвуя в формировании споровых покровов и кортекса. Третьи допускают, что выросты на спорах - результат каких-то нарушений в нормальном обмене веществ.
Химические анализы показали, что выросты состоят в основном из белка. В веществе выростов обнаружено 18 различных аминокислот, а также полисахариды и липиды. Оказалось, что по аминокислотному составу и структуре белок выростов сходен с такими белками, как кератин, актин, коллаген. Для того чтобы проделать подобные анализы, нужно было "отрезать" выросты у большого количества спор, а затем очистить от загрязняющих веществ и собрать их вместе. В качестве "ножниц" при этом был использован ультразвук, а собрать микроскопические отдельные отростки удалось с помощью современных высокоскоростных центрифуг.
Очень важным является вопрос о ферментативной активности выростов. В трубчатых выростах Bacillus penicillus выявлена дегидрогеназная активность (т. е. активность ферментов, отщепляющих водород от окисляемых субстратов).
Химический состав и тонкая структура выростов, их локализация в области контактирующих мембран двух функционально различных клеток (материнской клетки и проспоры) позволяют в общем виде заключить, что выросты бактериальных спор представляют собой специализированные структуры, которые ответственны за контакт клеток в системе спорангий - спорта.
Колпачки на спорах. Еще одной чрезвычайно интересной особенностью спорообразования у анаэробов является формирование на спорах блестящих колпачков. Эти колпачки на спорах у клостридиального и бациллярного типа клеток имеют конусовидную, а на спорах плектридиев - серповидную форму (рис. 48 - 50, 62). Колпачки при наблюдении в фазово-контрастном микроскопе четко выделяются благодаря сильному блеску. Электронная микроскопия показывает, что они имеют ячеистое строение. Множество отдельных ячеек упакованы таким образом на споре, что образуют сотовидную структуру в виде конуса или серпа. Ячейки - это электронно-прозрачные мешочки, заполненные газообразным веществом (газовые вакуоли). Мешочки имеют палочковидную или яйцевидную форму (см. рис. 62). Назначение колпачков состоит, по-видимому, в том, чтобы увеличивать подвижность и плавучесть спор в водной среде, и тем самым способствовать их распространению в природе. Подобно понтонам ячеистые колпачки не позволяют спорам тонуть. Споры с колпачками не удается осадить даже с помощью центрифуг, более того, при центрифугировании они всплывают на поверхность.
Способностью образовывать ячеистые колпачки обладают азотфиксирующие сахаролитические клостридии, сульфатредуцирующие бактерии и специфические почвенные плектридии, распространенные главным образом в почвах гидроморфного ряда.
Физиология спорообразования у анаэробов. Причины, вызывающие переход бактерий от вегетативного развития к спорообразованию, длительное время были неизвестны микробиологам. Впервые в нашей стране А. Я. Мантейфель, Н. Д. Иерусалимский провели исследования, вскрывшие факторы, влияющие на образование спор сахаролитическими анаэробами. Оказалось, что спорообразование вызывается исчезновением из среды источников азота, а также накоплением продуктов обмена (например, бутилового спирта). Но чтобы приобрести способность к споруляции, бактерии должны развиваться при нормальных условиях на полноценной питательной среде, содержащей набор аминокислот и факторов роста. Кислотность среды, оптимальная для вегетативного роста, является наиболее благоприятной и для спорообразования (рН 4,5 - 5,5 для CI. acetobutylicum и ~7 - 7,2 для протеолити-ческих анаэробов). Аэрация останавливает споруляцию. Эти данные свидетельствуют о том, что спорообразование является нормальной стадией развития у этих бактерий.
У анаэробов в отличие от аэробных видов исчерпание в среде источников углерода не влияет на процесс спорообразования. Даже добавление к среде глюкозы (5 г на литр) не сказывается на процессе спорообразования. Если культура выращена на полноценной питательной среде и приступила к спорообразованию, то дальнейший перевод клеток в условия полного голодания (например, перенос в водопроводную воду) не только не подавляет, но даже стимулирует споруляцию у некоторых сахаролитических анаэробов (CI. saccharobutyricum), т. е. для спорообразования не требуется в этом случае поступления питательных веществ из внешней среды (экзогенных источников питания), а для завершения конечных этапов формирования спор используются внутриклеточные запасы. Такой обмен веществ называется эндогенным (или эндотрофным) метаболизмом. Действительно, установлено, например, что белок спор образуется за счет азота белка вегетативных клеток. Однако у протеолитических анаэробов споруляция не поддерживается в среде, полностью лишенной органических питательных веществ. Так, для спорообразования CI. botulinum требуются аминокислоты - аланин и аргинин, которые используются в качестве источников энергии.
Генетический контроль спорообразования. Процесс "поглощения" материнской клеткой от сеченной на полюсе части цитоплазмы с ядерным элементом приводит к образованию новой клетки с двумя мембранами (проспора). Возникает особая ассоциация клеток - двуклеточный организм в общей оболочке, причем одна из клеток - проспора осуществляет свою жизнедеятельность внутри цитоплазмы другой, материнской клетки. Возникает крайне интересная ситуация, где сталкивается деятельность двух генетических центров - ядра материнской клетки и ядра споры. Многие стороны в их взаимоотношениях остаются еще неясными. Но из цитологических, физиологических и генетических наблюдений можно сделать вывод, что эти два ядра "разделяют" между собой обязанности, связанные с контролем над процессами формирования споровых структур.
Так, синтез новых белковых веществ, споровых оболочек, кортекса и экзоспориума осуществляется при активном контроле генома материнской клетки. При этом роль материнской клетки сводится к тому, чтобы как-то отделить от себя дочернюю клетку, оказавшуюся внутри ее цитоплазмы. Но так как удалить проспору за пределы клеточных границ через цитоплазматическую мембрану и клеточную стенку невозможно, то материнская клетка поступает с дочерней по-иному - она изолирует ее от себя путем инцистирования, окружая проспору мощными слоями белковой оболочки и переводя ее в метаболически неактивное (анабиотическое) состояние. Но это все равно не спасает материнскую клетку от гибели. Хотя она и продолжает еще длительное время активно функционировать, даже иногда способна образовать еще одну спору, но все же она не в состоянии вернуться к вегетативному развитию. В результате нарушения обмена веществ в конце концов наступает лизис (разрушение) вегетативной материнской клетки и освобождение спор.
Участие в спорогенезе большого количества ферментов и антигенов, включение новых путей биосинтеза и появление новых специфических клеточных структур показывает, что число генов, контролирующих спорогенез, достаточно велико (по всей вероятности, их больше 100). Некоторые из этих генов функционируют и в период вегетативной стадии развития. До сих пор остается неясным механизм функционирования спорового генома (набора генов, ответственных за спорообразование). Установлено, что споры получают весь свой ядерный материал (ДНК) в готовом виде от материнских клеток. При этом количество ДНК на спору постоянно для данного вида и не зависит от среды выращивания бактерий. Таким образом, этот показатель, вероятно, может служить характеристикой вида и использоваться при систематике спорообразующих бактерий.
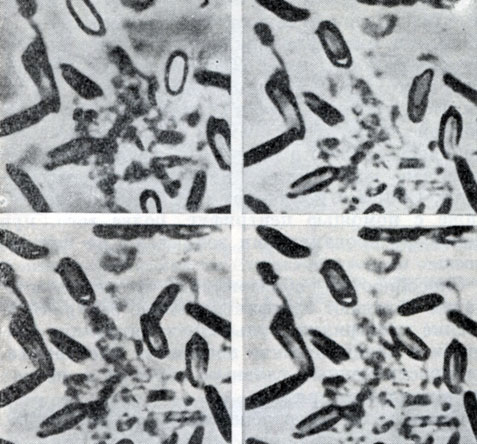
Рис. 63. Стадии прорастания спор CI. sporopenitum. Увел. X 2100
Прорастание спор. Споры, перенесенные в свежую питательную среду, начинают прорастать. Сначала они набухают, темнеют, затем через образовавшееся отверстие в оболочке споры молодая клетка выходит в среду. При этом слой кортекса разрушается, а споровая оболочка вместе с отростками (если таковые имеются) сбрасывается. У анаэробов проследить за прорастанием, наблюдая за одной и той же спорой, удается только в редких случаях. На рисунке 63 представлена серия фотографий, иллюстрирующих различные стадии прорастания - от потемнения сердцевины спор до выхода молодой вегетативной клетки из споровой оболочки. На последней стадии выхода вегетативная клетка как бы отстреливается. Отверстие в оболочке споры образуется не строго на полюсе споры, а несколько сбоку, и молодая вегетативная клетка при выходе расположена под углом к длинной оси споры. У других анаэробов процесс прорастания может выглядеть иначе.
Особенности прорастания GI. pasteurianum С. Н. Виноградский (1902) использовал для дифференциации этого вида от других спорообразующих анаэробов. Наконец, три вида клостридиев - CI. pectinovorum, CI. butyricum и CI. tetani - отличаются тем, что прорастание их спор происходит внутри спорангия. Клеточная стенка (или часть ее) у этих видов не лизируется, а остается на зрелых спорах, облегая их в виде чехла. Но этот чехол не идентичен по происхождению и строению с экзоспориумом, описанным выше.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'