В недрах цветка
Итак, насекомое или птица, ветер или вода доставили пыльцевое зерно в цветок и оно прикрепилось к липкой поверхности рыльца. Произошло опыление. Но это лишь предпосылка к оплодотворению, которое должно последовать позже и на пути к которому мужскому гаметофиту еще предстоит проявить достаточную активность: пыльце нужно прорасти на рыльце, а пыльцевой трубке - достигнуть зародышевого мешка, то есть женского гаметофита.
Зрелое пыльцевое зерно - это округлое образование, включающее в себя так называемое вегетативное ядро, которое плавает в богатой крахмалом и жирами клеточной плазме, и генеративную клетку, плавающую в той же плазме. Пыльцевое зерно покрыто двумя оболочками - внешней и внутренней. Внешняя оболочка зерна пористая, и это обстоятельство играет существенную роль при оплодотворении.
Кстати говоря, по химическому составу пыльца очень богата питательными веществами. Так, например, у кукурузы она содержит 43% крахмала, 40% аминокислот и нуклеиновых кислот, 2% жиров. Недаром цветочную пыльцу так охотно потребляют и насекомые, и тропические птицы-опылители.
Попадая на рыльце цветка своего вида, пыльцевое зерно прорастает на нем, образуя пыльцевую трубку, по которой спермин добираются до яйцеклетки, покоящейся в зародышевом мешке. Прорастание начинается с того, что внутренняя оболочка пыльцевого зерна внедряется в одну из пор внешней и разрастается в виде сосочка, в который как бы перекачивается цитоплазма зерна, его вегетативное ядро и генеративная клетка. Первым обычно проникает в пыльцевую трубку вегетативное ядро.
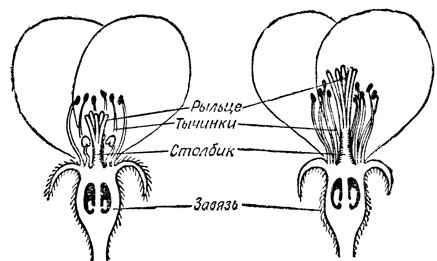
Рис. 24. Почему во многих обоеполых цветках никогда не происходит самоопыления? Один из механизмов такой защиты можно увидеть в цветке яблони. Сначала тычинки, щедро снабжающие пыльцой насекомых, возвышаются над несозревшим рыльцем. Тем временем столбик растет, и к моменту, когда пятилопастное рыльце становится способным принять пыльцу, оно возвышается над тычинками
Вскоре после проникновения в пыльцевую трубку генеративная клетка делится (митотически) и образует два спермия. Каждый из них представляет собой микроскопически малую клеточку, которая состоит из ядра, окруженного густой, однородной на вид цитоплазмой. Иногда спермин формируются в зрелом пыльцевом зерне еще до выхода генеративной клетки в пыльцевую трубку.
Пыльцевая трубка сначала внедряется в ткань рыльца, а затем проникает в столбик пестика. Растущий конец ее выделяет вещества, способные растворять межклеточные ткани, и пыльцевая трубка все глубже и глубже внедряется между клетками столбика, направляясь внутрь завязи - к семяпочкам. Любопытна и еще не объяснена эта способность трубки расти именно в нужном направлении, кратчайшим путем выходя к зародышевому мешку. Здесь не исключены какие-то химические влияния, помогающие трубке находить нужный путь.
Минимальный промежуток времени, необходимый для достижения пыльцевой трубкой семяпочки, равен 15 мин. Чаще всего этот срок составляет несколько часов. Но иногда процесс роста пыльцевой трубки растягивается на несколько месяцев. Так, у нашей березы он длится целый месяц, у ольхи и лещины - лесного ореха - от месяца до двух, а у некоторых представителей тропических орхидей (у них к моменту опыления семяпочки еще не достигают полной зрелости) - даже 6 - 7 месяцев. В последнем случае долгое прорастание, по-видимому, связано с сезонными явлениями, которым подчинена жизнь этих орхидей. Получается так, что опыление у них происходит в один дождливый сезон, а распространение семян - в следующий. Цикл оплодотворения и развития семян растянут таким образом на целый год - от одного дождливого сезона до следующего. Длительность прорастания пыльцы у наших древесных пород вызвана сходными причинами, ведь они вынуждены спешить с цветением: их цветки опыляются ветром и, как уже об этом говорилось выше, целесообразно, чтобы опыление произошло до распускания листьев. Для развития семян нужны поступающие от материнского растения питательные вещества, которые образуются уже летом, когда дерево во всю силу развернет работу своих "фабрик" органического вещества - зеленых листьев.
Любопытно, что у одного и того же вида растений рост пыльцевой трубки бывает быстрым в случае, когда происходит перекрестное опыление, и замедленным при самоопылении. Очевидно, в тех возможных случаях, когда на рыльце одновременно попадут пыльцевые зерна своего и чужого цветков, эта разница в скорости прорастания способствует тому, чтобы произошло более желательное для организма перекрестное опыление.
Достигнув завязи, пыльцевые трубки растут вдоль внутренней ее стенки и попадают в семяпочки через микропиле - пыльцевход. Это - наиболее типичный случай. Но иногда пыльцевая трубка проникает сначала в стенку завязи и только потом в семяпочку через ее основание. Так растут пыльцевые трубки в цветках березы, ольхи, лещины, граба и грецкого ореха.
По пыльцевходу пыльцевая трубка дорастает до вершины зародышевого мешка, растворяет его оболочку и изливает в него свое содержимое: цитоплазму, вегетативное ядро и оба спермия. Последние обычно проходят свой путь в зародышевом мешке через содержимое одной из синергид. К этому времени спермин теряют свою цитоплазму и имеют вид голых ядер.
О дальнейшей судьбе спермиев следует рассказать особо.
Где-то в начале книги нам пришлось упомянуть о борьбе за признание того факта, что у растений есть пол. Борьба эта была достаточно долгой и завершилась лишь менее 90 лет назад. Окончательно и воочию половую природу покрытосеменных растений доказал талантливый немецкий исследователь Э. Страсбургер в 1884 г. На примере цветка подъельника он показал все последовательные фазы внедрения пыльцевой трубки (мужского гаметофита) в зародышевый мешок (женский гаметофит), а затем и слияние ядра спермия с ядром яйцеклетки.
Но в пыльцевой трубке, как уже было сказано, образуется не один, а два спермия. Э. Страсбургер проследил судьбу одного. Лишь 14 лет спустя, в 1898 г., русский ботаник, эмбриолог С. Г. Навашин установил, что у покрытосеменных растений при оплодотворении один из спермиев (точнее, ядро его) соединяется с ядром яйцеклетки, а другой (опять-таки лишь ядро его) - со слившимися перед этим ядрами центральной клетки зародышевого мешка. Этот процесс получил с той поры название двойного оплодотворения.
В итоге слияния спермия с ядром яйцеклетки возникает зародыш, а второй спермий с ядрами центральной клетки дают начало эндосперму - особой питательной ткани, которую в последующем развитии использует зародыш.
Так происходит оплодотворение. И моментом его, моментом слияния спермиев с ядром яйцеклетки и ядрами центральной клетки, кончается невидимая непосвященным жизнь поколения мужского и женского гаметофитов - "мамы" и "папы" проростка, которому еще предстоит взойти из семени.
Впрочем, третье, "внучатое" поколение не спешит покинуть растение-"бабушку". Зародыш его еще проходит период созревания. Но об этом будет рассказано в следующей главе. А здесь следует упомянуть об отклонениях от классического двойного оплодотворения - отклонениях, на которые, как мы уже неоднократно имели возможность убедиться, очень щедра природа.
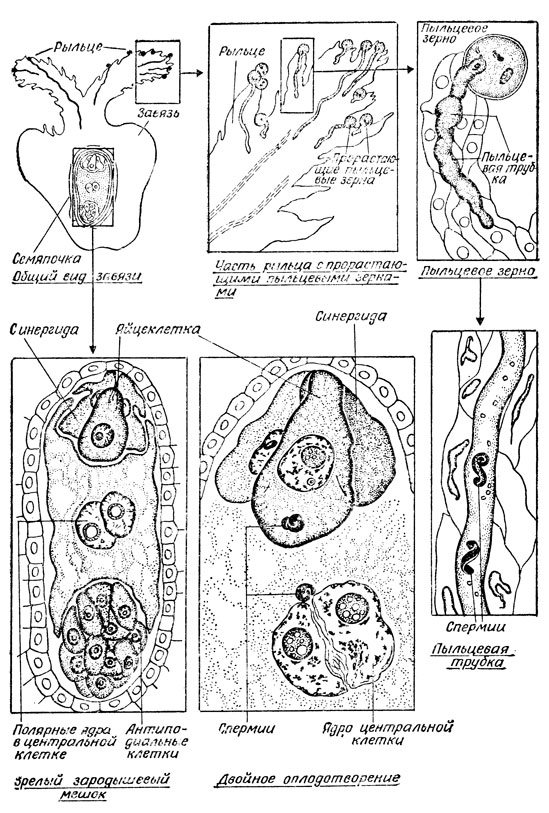
Рис. 25. Двойное оплодотворение у пшеницы. Этот важный этап знаменует конец раздельной жизни мужского и женского гаметофитных поколений и начало поколения спорофитного
Почти в то же время, когда С. Г. Навашин открыл двойное оплодотворение у растений, шведские ботаники Г. Юэль и С. Мурбек установили, что у некоторых растений яйцеклетка способна развиваться в зародыш вообще без оплодотворения - без слияния со спермием. Так был открыт партеногенез у растений - возможность развития зародышей и семян из девственной яйцеклетки, без участия мужской половой клетки.
Еще позже было установлено, что у некоторых видов орхидей происходит слияние спермия только с ядром яйцеклетки.
Наконец, возможно - и наблюдалось исследователями - излияние в зародышевый мешок содержимого не одной, а нескольких пыльцевых трубок.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'