Порядок аронниковые (Arales)
Семейство аронниковые, или ароидные (Araceae) (И. А. Груздинская)
Аронниковые - одно из крупных семейств однодольных, включающее около 110 родов и более 1800 видов, распространенных главным образом в тропических и субтропических областях обоих полушарий. Много аронниковых и в умеренных областях, а некоторые из них заходят даже в субарктические районы, однако видовое и родовое разнообразие их вне тропиков невелико (менее 10% видов).
Представители семейства - наземные, болотные, редко водные травы с клубнями или более или менее удлиненными корневищами.
В тропических странах аронниковые часто достигают гигантских размеров. Стебли тропических аронниковых нередко древовидные, но вторичный рост у них отсутствует. Немало среди них также лиан и эпифитов. Ветвление стеблей аронниковых обычно симподиальное, редко моноподиальное. У большинства прямостоячих форм, даже у гигантских трав, нет надземных вегетативных стеблей, которые заменены клубнями и корневищами. Однако у лазающих растений настолько длинные надземные стебли, что они уже не могут поддерживать себя в вертикальном положении. Обычно они опираются на деревья и удерживаются на них с помощью воздушных придаточных корней-прицепок. Эти корни не проявляют геотропизма, отрицательно гелиотропичны и очень чувствительны к контактному раздражению. Они отходят от повернутой к дереву-опоре стороны стебля, растут горизонтально, достигая иногда значительной длины, и "прилипают" к коре дерева-опоры с помощью особых волосков. Не менее обычны у аронниковых и питающие воздушные корни. Они более мощные и, в противоположность предыдущим, возникают на свободной, не прижатой к опоре стороне стебля. Эти корни растут вертикально вниз и свободно висят или как бы ползут вниз по коре дерева-опоры. Наконец, они достигают почвы, проникают в нее и интенсивно ветвятся, увеличивая активную всасывающую поверхность, и тем самым помогают обеспечить растение влагой и минеральным питанием. Добывают влагу воздушные питающие корни и другим путем. Их поверхность покрыта своеобразной, обычно многослойной покровной тканью - веламеном - и через ее мертвые клетки капиллярным путем, подобно губке, всасывается конденсирующаяся атмосферная влага. В корнях аронниковых обычны сосуды с лестничной перфорацией, но в стеблях они встречаются крайне редко, и водопроводящие элементы представлены главным образом трахеидами. Листья аронниковых очередные, в большинстве случаев расчлененные на черешок и пластинку, приземные или стеблевые, разных размеров и строения. Поражает чрезвычайное разнообразие листовой пластинки, но преобладают простые цельные широкие пластинки с сетчатым жилкованием. Однако есть все переходы к гигантским листьям со сложно рассеченными пластинками и мощными черешками. У примитивных членов семейства листья типичные для однодольных: узкие, длинные, с параллельным жилкованием, влагалищные и без черешков. Форма и рассеченность пластинки листа нередко поразительно меняются в процессе жизни растения. У многих аронниковых рассеченные крупные листья взрослых растений существенно отличаются от мелких цельных листьев их ювенильных побегов, как это хорошо видно у монстеры тонкой (Monstera tenuis, рис. 266). Почти полностью редуцирована пластинка листа и у висящих побегов столонов (рис. 267). Строение и форма пластинки могут существенно меняться в течение собственной жизни листа, и это особенно четко прослеживается при формировании своеобразнейших продырявленных листьев, характерных для многих аронниковых. У монстеры деликатесной (М. deliciosa), например, зачаток пластинки листа сначала цельный, и лишь с определенной стадии развития наблюдается сначала посветление отдельных участков, затем отмирание мезофилла, а потом и эпидермы, в результате образуются щели между некоторыми боковыми жилками. В дальнейшем эти щели увеличиваются, появляются новые, и, наконец, вдоль боковых жилок образуются крупные полости, доходящие до края пластинки листа. Затем разрывается край, и лист становится не только продырявленным, но и рассеченным. Иначе возникает рассеченность листа у филодендрона радиального (Philodendron radiatum). Как показал японский ботаник М. Хотта (1971), краевые участки зубчатого молодого листа его растут с неодинаковой скоростью: область боковой жилки развивается быстрее, чем пространство между жилками. В результате неравномерного роста сегменты с боковыми жилками оказываются глубоко разделенными и возникает пальчато-лопастный лист.
Разнообразно у аронниковых и строение черешков. В морфологическом ряду изменений от бесчерешковых листьев с влагалищами к черешковым без влагалища есть много промежуточных форм, у которых черешок выполняет также функцию влагалища и соответственно имеет двойственную структуру: с внешней стороны он выглядит типичным черешком, а с внутренней - типичным влагалищем. Характерны у аронниковых и другие видоизменения черешков. У эпифита филодендрона толстого (P. crassum) черешок служит хранилищем влаги, он сильно разрастается в толщину, становится водянистым, желтоватым и напоминает суккулентный стебель (рис. 260). У многих видов потоса (Pothos) черешки, напротив, совершенно плоские и имеют вид листовой пластинки (рис. 263). На очень тесную связь между формой черешка и его приспособлением к изменению ориентации пластинки листа по отношению к свету обратил внимание М. Хотта. Круглые черешки ряда филодендронов реагируют на свет перекручиванием, крылатые черешки некоторых монстер и потосов - сгибанием. При этом черешок сгибается обычно в верхней части в области небольшого утолщения, после которого ось черешка несколько смещается и образуется как бы коленный изгиб (рис. 263, 266).
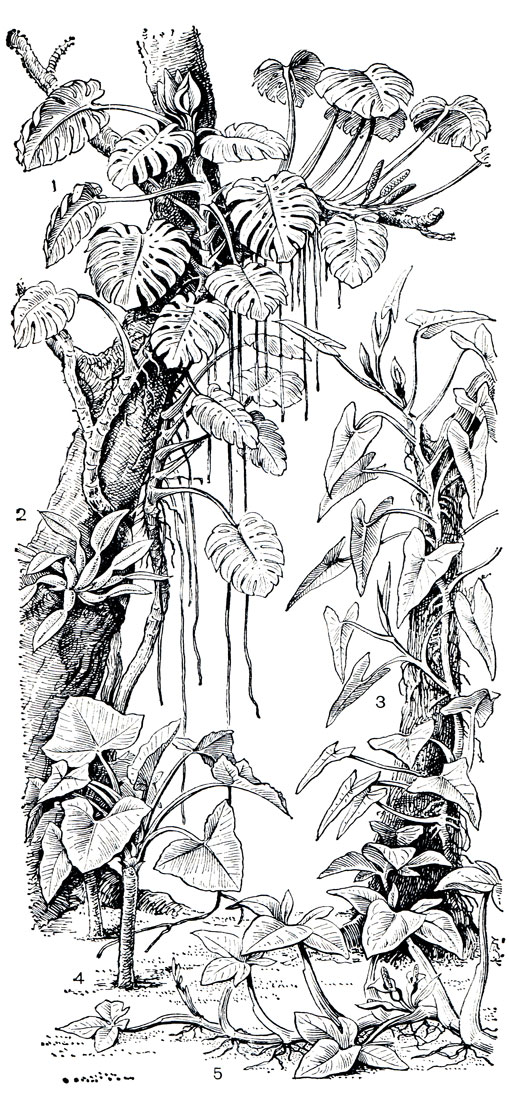
Рис. 260. Аронниковые тропических лесов Нового Света: 1 - монстера деликатесная (Monstera deliciosa); 2 - филодендрон толстый (Philodendron crassum); 3 - филодендрон имбе (P. imbe); 4 - ксантосома Жакэна (Xanthosoma jacquinii); 5 - сингониум ножколистный (Syngonium podophyllum)
У аронниковых обильны и разнообразны элементы выделительных тканей. Это отдельные выделительные клетки - идиобласты - с одиночными кристаллами оксалата кальция, друзами, рафидами, секреторные клетки, межклеточные трихосклереиды, а также схизогенные вместилища, смоляные каналы и особенно часто встречающиеся членистые млечники. Помимо выделительной функции многие из этих образований играют роль защиты растения от поедания животными.
У аронниковых лишь один тип соцветия - початок, на котором обычно очень плотно, сериями спиралей размещены лишенные прицветников мелкие невзрачные цветки, которые не всегда можно четко разграничить. Цветки обоеполые или однополые; обоеполые цветки в большинстве случаев с 4-6-членным околоцветником, реже голые; однополые цветки обычно голые и лишь как исключение с околоцветником. Тычинок 4-6, но их число может быть редуцировано до 1 или возрасти до 8. Тычинки свободны или срастаются в специфические образования - синандрии. У специализированных групп нити тычинок редуцированы и непомерно разросшийся связник превращает тычинки в подобие геометрических фигур: призм, квадратов, усеченных пирамид. Пыльники яйцевидные или линейно-продолговатые, раскрывающиеся порами, продольными или поперечными щелями. Пыльцевые зерна с оболочкой разнообразных типов. Гинецей ценокарпный из 2-3 (до 9) плодолистиков, иногда псевдомономерный; завязь верхняя, лишь иногда погруженная в мясистую ось соцветия, 1-3-гнездная, с одним или многими семязачатками в каждом гнезде. Зародыш с обильным эндоспермом или иногда без него. Плод почти
у всех аронниковых - одно многосемянная ягода, обычно ярко окрашенная.
У аронниковых цветение проходит в 2 фазы. Сначала функционируют рыльца (женская фаза цветения), и только после потери ими способности воспринимать пыльцу раскрываются пыльники (мужская фаза цветения). Протогиния встречается у растений с обоеполыми и с однополыми однодомными цветками. Последовательность женской и мужской фаз цветения препятствует самоопылению, однако у аронниковых это достигается далеко не всегда. В женскую фазу вступают сначала нижние цветки, и их зацветание идет обычно по початку строго снизу вверх. Развитие же и раскрывание пыльников, как правило, не имеет такой последовательности. Нередко у верхних цветков или даже у цветков в средней части початка фазы цветения совпадают и становится возможным самоопыление. Это относится и к гейтоногамии, которая довольно обычна у аронниковых. Таким образом, протогиния не так уж надежно защищает цветки аронниковых от самоопыления, и для предотвращения его потребовалось развитие других механизмов. Одним из них становится структура самого соцветия. Эволюция соцветий в семействе идет по пути все более резкого отграничения женских частей цветка и соцветия от мужских. И если у примитивных аронниковых цветки обоеполые, то в конце эволюционного ряда у наиболее специализированных групп цветки однополые, причем женские и мужские размещены в разных частях соцветия или даже на разных растениях (двудомные цветки у ряда аризем - Arisaema). Самоопыление в этом случае совершенно исключено (рис. 261).
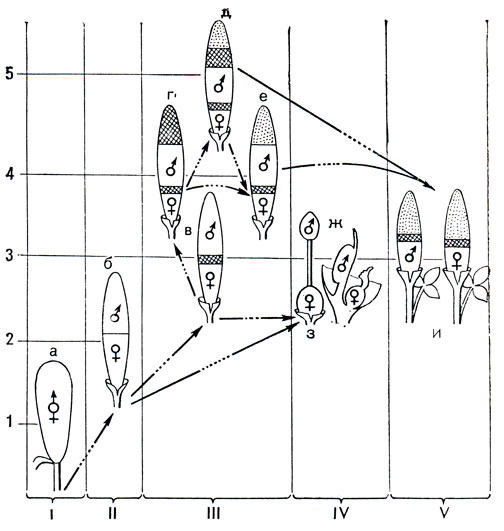
Рис. 261. Схема предполагаемой эволюции соцветий у аронниковых. I - V - специализация соцветий в направлении разобщения пола (от обоеполых до двудомных цветков), 1-5 - усложнение структуры соцветия (увеличение числа зон на початке). Зоны цветков: ♀ - женских; ♂ - мужских; комбинированный значок - обоеполых. Заштрихована зона стерильных цветков (верхняя и нижняя), точками показан придаток соцветия, иногда превращающийся в осмофор (орган запаха). Аирные: а - аир обыкновенный (Acorus calamus). Лазиевые: б - анхоманес двухформный (Anchomanes diformis). Филодендровые: в - филодендрон Варшевича (Philodendron warszcewiczii). Колоказиевые: г - схизматоглоттис коротконожковый (Schismatoglottis brevipes); е - алоказия ладьевидная (Alocasia navicularis). Собственно аронниковые: д - аронник пятнистый (Arum maculatum); ж - амброзиния Басса (Ambrosinia bassii); з - стилохитон ланцетолистный (Stylochiton lancifolius); и - аризема амурская (Arisaema amurense): разнополые цветки расположены на разных растениях (двудомность)
Соцветия аронниковых поражают разнообразием и за редкими исключениями выглядят как одиночные цветки. Такое впечатление создается главным образом благодаря видоизменению покрывала (кроющего листа) соцветия, нередко ярко окрашенного и принимающего форму околоцветника. Иногда она так причудлива, что соцветие можно принять за экзотический цветок орхидных или лист-кувшинчик насекомоядных непентесов. Отдельным цветком выглядит и огромное соцветие аморфофаллуса гигантского (Amorphophallus titanum), достигающее в высоту 2-3 м и развивающее до 5000 цветков, и 3-9-цветковое соцветие пистии телорезовидной, или водного латука (Pistia stratiotes),- крошечное, умещающееся в сантиметре. Но отличия не только в размерах, соцветия различаются и по многим существенным признакам, показывающим разную степень их эволюционной продвинутости. Наиболее примитивные соцветия, несущие на початке только обоеполые цветки, свойственны самым примитивным аронниковым (рис. 261, 262). В более специализированных подсемействах развиваются только однополые цветки. На початке образуются две зоны цветков: нижняя - из женских цветков, верхняя - из мужских (рис. 261). Затем в области их контакта возникает зона стерильных цветков, а иногда образуется даже вторая зона стерильных цветков на верхушке початка. В некоторых случаях стерильные цветки редуцируются, и тогда женскую часть соцветия от мужской отделяет лишь голый участок стерильной оси соцветия (рис. 261). У некоторых аронниковых верхние стерильные цветки образуют придаток початка, нередко превращающийся в так называемый осмофор - носитель запаха, привлекающего опылителей. У отдельных видов этот придаток приобретает причудливую форму шляпки гриба или становится нитевидным (рис. 277). Видоизмененные стерильные цветки также играют определенную роль в опылении насекомыми, особенно у специализированных аронниковых. В пределах семейства параллельно усложнению строения соцветия упрощалось строение цветков, и в специализированных подсемействах женский цветок состоит обычно только из одного гинецея, а мужской часто сведен к 1 тычинке или 1 синандрию. Нередко мужские цветки на одном соцветии состоят из разного числа тычинок (как, например, у аморфофаллусов), и тогда определить границу отдельного цветка очень трудно.
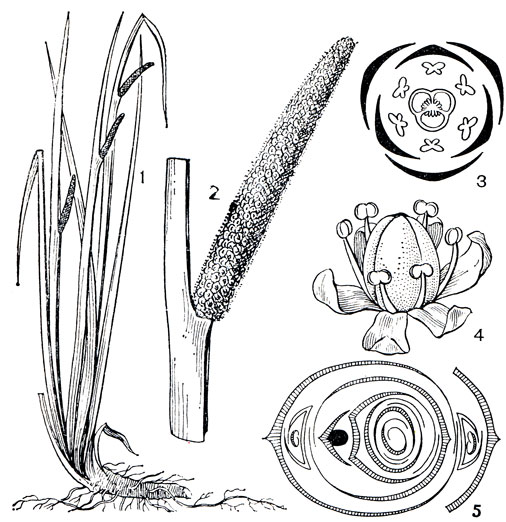
Рис. 262. Аир обыкновенный (Acorus calamus): 1 - общий вид; 2 - соцветие-початок; 3 - диаграмма цветка; 4 - цветок; 5 - диаграмма растения
В связи со специализацией соцветия интересны и изменения покрывала соцветия. У аирных оно не отличается от обычного листа и быстро опадает. Но у большинства специализированных групп покрывало полностью или частично прикрывает соцветие, выполняя, помимо защитной, и другие функции. Свойственная многим видам яркая окраска покрывала привлекает насекомых-опылителей, особое строение покрывала соцветий-ловушек способствует "ловле" насекомых-опылителей и удержанию их в соцветии в зоне расположения женских цветков. Двухкамерное покрывало у видов криптокорин (Cryptocoryne) препятствует самоопылению и предохраняет соцветие от намокания (рис. 279).
Опыляются цветки аронниковых преимущественно насекомыми (мухами, пчелами, жуками, тлями). Некоторое значение в опылении, хотя и оспариваемое отдельными исследователями, имеют улитки, в ряде случаев не исключено и ветроопыление. Для аронниковых характерен особый род энтомофилии - сапромиофилия - опыление навозными и падальными мухами. В этом случае растение оказывается активным членом, оно как бы заставляет насекомых против их воли опылять цветки. Для этого у растения формируется ряд специальных структур, предназначенных для обмана насекомого. Имитируя запах и цвет субстрата, в который откладывают яйца эти насекомые, растение буквально зазывает опылителей в соцветие-ловушку и держит их пленниками до тех пор, пока они не опылят цветки и не получат пыльцу для опыления цветков других соцветий. У сапромиофильных аронниковых цветение сопровождается совершенно необычным для высших растений явлением, описанным еще Ламарком более 200 лет назад, но и до сих пор не переставшим интересовать ученых. Это резкое повышение температуры соцветия или отдельных его частей на 10, 16 и даже 30 °С по сравнению с температурой окружающей среды. Но особенно замечательна тесная связь между быстрым повышением температуры и столь же быстрым появлением от початка крайне неприятного запаха. Оба эти явления непродолжительны и обычно исчезают через несколько часов. Последующие исследования установили, что появление запаха сочетается с огромной метаболической активностью в соцветии и связано с чрезмерной активностью дыхания, что само по себе может привести к повышению температуры. Под воздействием тепла начинают испаряться летучие вещества, несущие запах, и распространяющееся зловоние привлекает мух-опылителей. Хроматографическое исследование веществ, составляющих запах початка аронниковых, обнаружило еще одно интересное звено в цепи рассматриваемых явлений - необычайно быстрое, взрывное увеличение количества свободных аминокислот в тканях соцветия во время раскрывания пыльников. В початке сауроматума капельного (Sauromatum guttatum), как показал голландский исследователь А. ван Херк (1937), концентрация свободных аминокислот в период цветения буквально за один день повысилась в 20 раз. Пока даны лишь самые общие объяснения всем этим явлениям, и в настоящее время их исследование перешло на субмикроскопический уровень, в область изучения деятельности митохондрий.
Неприятный запах, исходящий от соцветий аронниковых, иногда бывает связан с другим типом опыления - сапрокантарофилией, как, например, у аморфофаллуса гигантского. Отвратительный запах соцветия привлекает навозных и падальных жуков - его постоянных опылителей. Многие аронниковые из разных групп родства в качестве опылителей используют пчел, ос, тлей и привлекают их приятным цветочным запахом и сладковатой жидкостью, похожей на нектар. Аронниковые не имеют морфологически выраженных нектарников, и сладковатая жидкость, выполняющая функцию нектара, выделяется у них разными органами цветка. По данным чешского исследователя Е. Дауманна (1931), эта жидкость у видов аронника (Arum) может скапливаться на волосках рылец, у аглаонемы (Aglaonema) - на тычинках, у антуриумов - на внешней поверхности околоцветника, куда попадает через устьица, у представителей подсемейства монстеровых - в стаминодиях женских цветков и на рудиментах гинецеев в мужских цветках, у ряда видов ариземы - на покрывале женских соцветий.
В пределах аронниковых выделяют обычно 9 подсемейств. Начиная от наиболее примитивного, они образуют следующий ряд нарастания специализации: аирные (Acoroideae), потосовые (Pothoideae), монстеровые (Monsteroideae), калловые (Calloideae), лазиевые (Lasioideae), филодендровые (Philodendroideae), колоказиевые (Colocasioideae), собственно аронниковые (Aroideae) и пистиевые (Pistioideae). В таком порядке мы их и рассмотрим.
Подсемейство аирных объединяет растения с узкими длинными листьями без черешков, с влагалищами и параллельным или почти параллельным жилкованием. Цилиндрические соцветия-початки несут одинаковые обоеполые 3(4)-членные цветки с околоцветником; тычинки свободные, гинецей из 3 плодолистиков, завязь 1-3-гнездная с ортотропным висячим семязачатком в каждом гнезде. Семена с эндоспермом. Кроющий лист соцветия узкий, невзрачный, не прикрывает початок. В этой небольшой группе из 2 родов и 3 видов совершенно необычные географические связи. Род гимностахис (Gymnostachys) с единственным видом гимностахис обоюдоострый (G. anceps) обитает в тропических лесах Восточной Австралии; род аир (Acorus) широко распространен во внетропических областях северного полушария. Аир обыкновенный (A. calamus) - хорошо знакомое нам небольшое травянистое растение заболоченных мест, обитающее вдоль рек, по берегам стариц, окраинам болот. От его горизонтально простирающихся корневищ отходят снизу корни, сверху листья, по форме сходные с листьями ириса, и цветоносные побеги (рис. 262). Все части этого растения издают едва уловимый приятный аромат. Цветет аир ранней весной, развивая на початке множество желтоватых цветков, однако цветение далеко не всегда оканчивается образованием плодов - зеленоватых суховатых ягод, распространяемых животными. Аир расселяется главным образом с помощью вегетативного размножения- корневищами. В распространении его на большие расстояния определенное значение приобретает вода. Оторванные куски корневищ долго могут плыть по течению реки, пока не прибьются к берегу и не укоренятся. Аир издавна широко культивируют даже в тропических странах, и уже более 4000 лет им торгуют на Ближнем Востоке. В XIII в. аир был ввезен в Польшу как лекарство, в XVI в. его стали разводить в Западной Европе. Внимание к этому растению объясняется высоким содержанием почти во всех его частях ароматичного эфирного масла, используемого в разнообразнейших целях. Благодаря этой особенности аир был распространен человеком по всему северному полушарию, где он успешно произрастает от европейской субарктики и атлантической Северной Америки до тропических островов Юго-Восточной Азии. Предполагаемой родиной этого растения считают Восточную Азию. В наши дни аир обыкновенный тоже используют как источник получения эфирного масла сложного химического состава, называемого аирным или ирным. Масло извлекают преимущественно из мясистых корневищ, содержащих его до 4,5%, и используют в парфюмерии, медицине, при изготовлении ликеров, вин и пива. В народной медицине порошком из высушенных корневищ лечат всевозможные заболевания. Молодые побеги используют вместо салата. Аир обыкновенный разводят и как декоративное водное растение. В этих целях используют также аир злаковый (A. gramineus) - небольшое узколистное растение, по внешнему облику очень похожее на злак.
К примитивным аронниковым относят и большое пантропическое подсемейство потосовых. Как и аирные, его представители не имеют млечников и развивают преимущественно обоеполые цветки. В подсемействе около 15 родов, в том числе и самый крупный в семействе неотропический род антуриум (Anthurium), насчитывающий около 500 видов. Жизнь потосовых тесно связана с тропическими лесами.
Антуриум представлен преимущественно травянистыми растениями со сравнительно толстыми, нередко укороченными стеблями и междоузлиями, реже стебли удлиненные, иногда ползающие или лазающие и очень редко древовидные. Листья разнообразной формы и рассечения (от цельных до сложно рассеченных),} черешки всегда с геникулумом. Покрывало соцветия чаще зеленое, но нередко и ярко окрашенное, подобно околоцветнику, в красный, фиолетовый, розовый цвета. Цветки антуриумов сериями спиралей тесно расположены на цилиндрическом початке (рис. 263) и похожи на ромбы и квадраты. Цветки всегда обоеполые, обычно с 4-членным околоцветником и 4 тычинками; гинецей синкарпный; рыльце чаще дисковидное; завязь 2-гнездная, с 1-2 семязачатками в каждом гнезде. Плоды - сочные мясистые ягоды; семена плосковыпуклые, с мясистым эндоспермом.
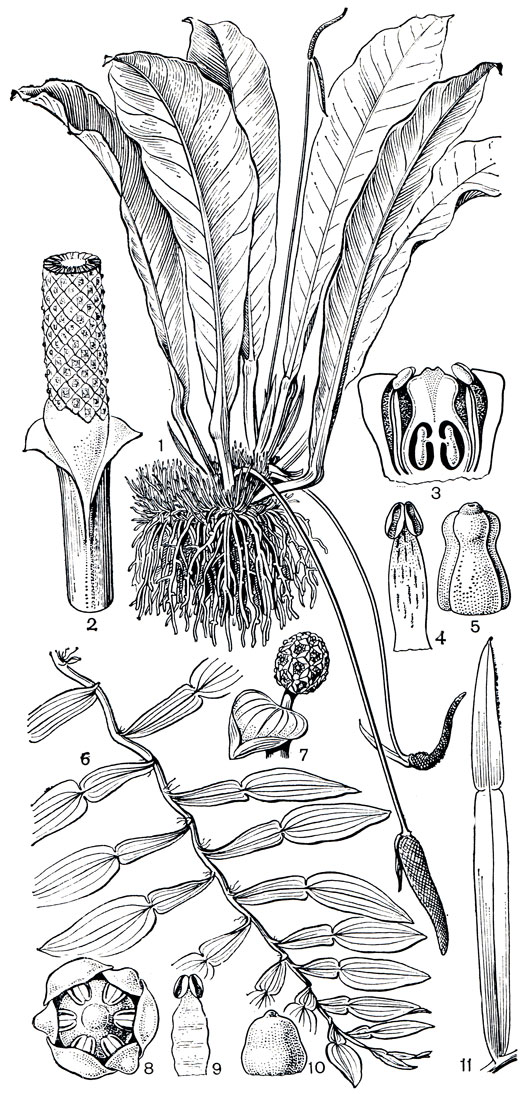
Рис. 263. Аронниковые: подсемейство потосовых. Антуриум толстожилковый (Anthurium crassinervium): 1 - общий вид; 2 - нижняя часть соцветия; 3 - продольный разрез цветка; 4 - тычинка; 5 - гинецей. Потос лазающий (Pothos scandens): 6 - лазающий побег. Потос Зееманна (P. seemannii): 7 - соцветие; 8 - цветок; 9 - тычинка; 10 - гинецей. Потос Лоурейры (P. loureirii): 11 - лист с плоским черешком
Цветение антуриумов своеобразно. Во время женской фазы в цветках видны только рыльца, тычинки же полностью скрыты в углублениях частей околоцветника (рис. 264) и цветок выглядит женским. На рыльцах в это время появляются капли сладковатой вязкой жидкости, привлекающие опылителей. После того как жидкость подсохнет и рыльце потеряет способность воспринимать пыльцу, из-под сегментов околоцветника появляются тычинки (рис. 264). Достигая полного развития, они у некоторых видов сильно вытягиваются и закрывают рыльце. Иногда же тычинки после опыления быстро втягиваются к основанию околоцветника, становятся незаметными, и цветок снова выглядит женским. У большинства антуриумов, как и у многих других аронниковых, цветки развиваются на початке от основания соцветия к его верхушке, но у некоторых первыми появляются цветки в центре початка, еще реже последовательность их появления установить не удается. Продолжительность цветения антуриумов, по наблюдениям американского исследователя Т. В. Кроата (1980), сильно колеблется у разных видов и может длиться от нескольких часов до нескольких недель. Сильно варьирует у разных видов и запах цветков, от едва ощутимого до очень сильного, приятного или зловонного, в зависимости от "запросов опылителя". Мух привлекают отврати-тельный запах и темная окраска початка, покрывала и листьев. Пчелы предпочитают сладкий, парфюмерный запах и светлые тона окраски. Т. В. Кроат считает, что иногда пчелы,, по аналогии с их поведением на орхидных, "собирают запах", а не пыльцу или нектар и используют запах растения для сексуального привлечения. "Надушившись", самцы с большим успехом привлекают самок. У цветков разных видов антуриумов запахи различаются по составу химических веществ и появляются в разное время суток. Цветки одних видов пахнут только утром, других - среди дня. Появление же неприятного запаха непосредственно не зависит от суточной периодичности и обычно связано с развитием тычинок. Среди опылителей цветков антуриумов - насекомые из разных и довольно далеких таксономических групп - пчелы, мухи, жуки и осы. Плод антуриумов - сочная, иногда ярко окрашенная ягода, разносимая обычно животными, преимущественно птицами. По мере созревания ягода как бы выдавливается из околоцветника и повисает на 2 нитевидных полосках, закрепленных на частях околоцветника. В этом более открытом положении на початке часть ягод какое-то время удерживается и становится более доступной для птиц.
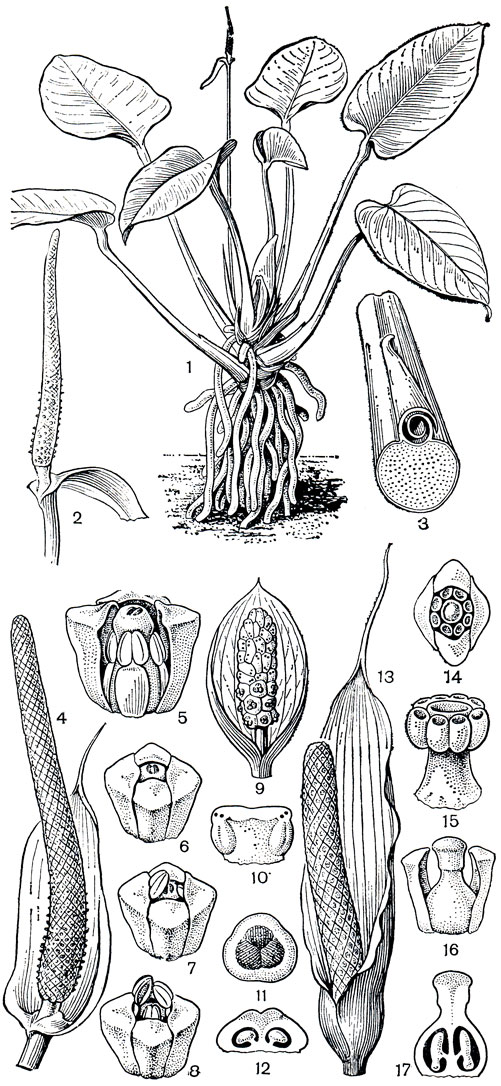
Рис. 264. Аронниковые: подсемейство потосовых. Антуриум (Anthurium sp.): 1 - общий вид, 2 - соцветие; 3 - поперечный разрез основания черешка со свернутым влагалищем. Антуриум изящный (A. elegans): 4 - соцветие в женской стадии цветения, видны капли вязкой жидкости на рыльцах нижних цветков; цветки: 5 - вид сбоку, передняя часть околоцветника удалена; 6 - в женской стадии цветения; 7, 8-начало мужской стадии цветения. Кульказия мелкополосатая (Culcasia striolata): 9 - соцветие; 10 - тычинка; 11 - гинецей (вид сверху); 12 - продольный разрез гинецея. Гонатопус Буавэна (Gonatopus boivinii): 13 - соцветие; 14 - мужской цветок (вид сверху); 15 - сросшиеся тычинки; 16 - женский цветок; 17 - продольный разрез гинецея
Антуриумы произрастают почти по всей тропической и субтропической Америке. Они широко распространены в Андах Южной и Кордильерах Центральной Америки почти от уровня океана до 3400 м над уровнем моря. Но даже у верхней границы леса (на высоте более 3000 м) антуриумы достигают довольно больших размеров. Многие высокогорные виды являются эпифитами. В нижнем поясе на высотах 100-300 м распространена своеобразная группа антуриумов, обитающих в саваннах. Надземный стебель у них редуцирован; грубая толстая пластинка листа сравнительно узкая, сильно вытянутая, черешок очень короткий; в основании растений формируются воздушные корневые гнезда с вверх направленными корневыми окончаниями, покрытыми веламеном. Это антуриум толстожилковый (A. crassinervium, рис. 263), антуриум эллиптический (А. ellipticum), антуриум морщинистый (A. rugosum) и др. Антуриумы широко используются как декоративные растения, и сейчас уже выведено много сортов. Всегда привлекают внимание антуриум Андрэ (A. andreanum) и антуриум Шерцера (A. scherzerianum) с ярко-красными покрывалами, сочетающимися с желтыми, оранжевыми или красными початками, нередко причудливо изогнутыми (табл. 61 и 63).

Таблица 61. Аронниковые: 1 - антуриум Шерцера (Anthurium scherzerianum), оранжереи Ботанического института АН СССР в Ленинграде; 2 - пистия телорезовидная, или водный латук (Pistia stratiotes), там же; 3 - антуриум овальнолистный (A. ovalifolium), там же; 4 - антуриум изменчивый (A. variabile), оранжереи Ленинградского государственного университета; 5 - алоказия Сандера (Alocasia sanderiana), оранжереи Ботанического института АН СССР в Ленинграде

Таблица 63. Аронниковые: 1 - антуриум Андрё (Anthurium andreanum); 2 - антуриум изящный (A. elegans); 3 - соплодие белокрыльника
В Старом Свете, преимущественно в муссонных областях, от Мадагаскара и Коморских островов, через тропическую Азию до архипелага Бисмарка и Восточной Австралии широко распространен типовой род подсемейства потос (Pothos), включающий примерно 75 видов. Виды потоса являются характерными растениями тропических лесов, многие из них - лазающие эпифиты со своеобразными плоскими черешками, очень похожими на листовые пластинки (рис. 263).
Своеобразный монотипный род замиокулъкас (Zamioculcas) распространен в горных каменистых степях Восточной Африки, где его вид замиокулъкас замиелистный (Z. zamiifolia) произрастает на открытых местах среди камней вместе с суккулентными молочаями и дорстениями; обитает он также и на прибрежных холмах Занзибара. Замиокулъкас обладает очень интересным вегетативным размножением. Каждый листочек его непарноперисто рассеченного листа опадает отдельно от общего черешка и в дальнейшем ведет себя как типичный выводковый лист. В основании опавшего листочка образуется клубневидное вздутие, развивающееся в настоящий клубень (рис. 265). Этот клубень укореняется и дает начало новому растению. Близкий к замиокулькасу род гонатопус (Gonatopus), включающий 2 вида, распространен в нижнем поясе горных лесов тропической Восточной Африки и по ряду признаков несколько отличается от типичных потосовых. Цветки у него однополые, в мужских цветках нити тычинок срастаются в трубку вокруг рудимента гинецея (рис. 264). Стебель подземный в виде сплюснутого клубня, развивающий множество корневищ. Единственный лист больше метра длины с пятнистым черешком и 3-4-перисторассеченной пластинкой.
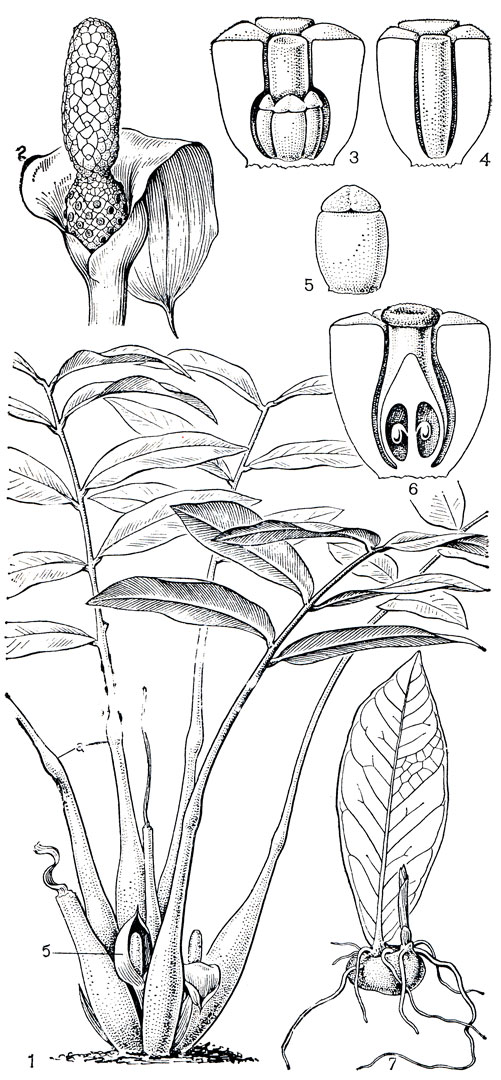
Рис. 265. Замиокулькас замиелистный (Zamioculcas zamiifolia): 1 - общий вид (а - генииулум, б - соцветие); 2 - соцветие с отогнутым покрывалом, нижние цветки женские, верхние - мужские; 3 - мужской цветок (передний сегмент околоцветника удален); 4 - стерильный цветок; 5 - тычинка; 6 - продольный разрез женского цветка; 7 - укоренившийся выводковый лист
Подсемейство монстеровых (Monsteroideae) - интереснейшая тропическая группа из 7 -10 родов и более 160 видов, преимущественно корнелазающих растений, обитателей тропических дождевых лесов и лесов полосы туманов. Монстеровые придают своеобразный облик этим лесам, обвивая ветви и стволы деревьев и прикрывая их крупными, нередко продырявленными, причудливо изрезанными листьями. Особенно хорошо они представлены в экваториальной Америке и Азии и лишь единичными видами - в тропической Западной Африке и в Австралии. В муссонных областях Старого Света распространен крупный род монстеровых рафидофора (Raphidophora), включающий более 60 видов. В тропиках Нового Света из крупных родов, помимо монстеры, произрастает также стеносперматион (Stenospermation), включающий более 20 видов прямостоячих растений, обычно с коротким надземным стеблем и цельнокрайными листьями. Некоторые виды его развиваются как настоящие эпифиты. Соцветие монстеровых закрыто покрывалом и раскрывается только ко времени цветения, после которого обычно опадает. Цветки у них обоеполые, но уже без околоцветника и лишь у наиболее примитивных родов, близких к потосовым, початок не завернут в покрывало - цветки имеют 2 круга околоцветника, нередко сросшегося. К этим исключениям принадлежит новогвинейский род голохламис (Holochlamys) и спатифиллум (Spathiphyllum, около 36 видов), ареал которого имеет огромный древний разрыв - тропическая Америка и Малезия. Плоды у монстеровых обычно сочные ягоды, тесно расположенные на початке и распространяемые в основном птицами. В тканях монстеровых нет млечников, в паренхиме образуются характерные трихосклереиды, которые называют также межклеточными волосками. Они возникают как небольшие треугольные образования в межклетниках, затем разрастаются и превращаются в длинные тонкие игольчатые образования с отростком в центре, иногда как бы соединяются парами в виде буквы Н.
Монстеровые часто называют полуэпифитами или теневыми эпифитами, хотя их семена, за исключением ряда видов стеносперматиона, произрастают наземно, и в течение всей последующей жизни растение не теряет связи с почвой. Имеется интересная особенность в развитии вегетативных лазающих побегов: когда растение достигает вершины ствола дерева-опоры или не может ползти дальше вверх, оно опять развивает плетевидные (столонообразные) побеги. В поисках новой опоры эти побеги горизонтально отклоняются, повисают и, находясь в таком состоянии, растут вниз, образуя очень длинные междоузлия (у монстеры остроконечной - Monstera acuminata - длиной до 30 см) и сильно редуцированные пластинки листьев (рис. 267). Наконец, побеги достигают почвы, укореняются и стелются по ее поверхности, пока не встретят опору, по которой с помощью корней-прицепок ползут вверх и превращаются в обычную лазающую лиану с крупными перисторассеченными листьями (рис. 266). Прямостоячие монстеровые (роды стеносперматион, спатифиллум, родоспата - Rhodospatha), в отличие от рассмотренных, образуют только один тип побегов.
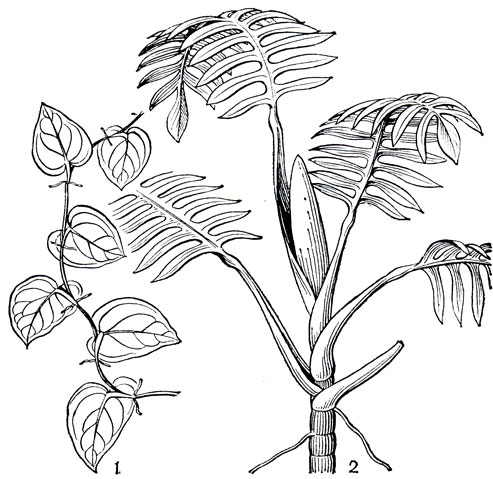
Рис. 266. Разнолистность у монстеры тонкой (Monstera tenuis): 1 - цельные листья ювенильного побега; 2 - перисторассеченные листья побега взрослого растения
Жителям умеренного климата наиболее знакомы из этого подсемейства представители рода монстера, широко культивируемые у нас в оранжереях, зимних садах, жилых помещениях. Название рода "монстера" (от monstrosus - ненормальный, причудливый) дано ему за необычные продырявленные листья (рис. 260, 267); до 40 отверстий бывает на крупных метровых листьях некоторых монстер. Монстера включает около 25 видов декоративных лазающих лиан, особенно широко распространенных в Центральной и Южной Америке.
Цветение монстер обычно не связано с годичной ритмикой, и их соцветия можно увидеть в любое время года. У многих видов перед цветением зеленоватое покрывало закрыто и полностью скрывает початок, и лишь к моменту, когда рыльца становятся способными принимать пыльцу, оно приоткрывается и его внутренняя поверхность окрашивается в желтоватые, беловатые или розовые тона. Наступает женская фаза цветения. Привлеченные ярким цветом покрывала и запахом ананаса, исходящим от соцветия, насекомые (небольшие мухи, пчелы, муравьи), в том числе и несущие пыльцу из других соцветий, проникают к початку и в поисках нектара ползают по соцветию, опыляя его цветки. Некоторые насекомые задерживаются в нижней части початка, где поглощают капли сладковатой жидкости, выделяющейся на рыльцах расположенных там стерильных цветков. Они выползают из соцветия позже, обычно в период мужской фазы цветения. Ей предшествует удлинение нитей тычинок и появление пыльников, до этого скрытых в углублениях шестигранного гинецея. Пыльники у монстер раскрываются порами, и "выдавливающиеся" из них липкие, волокнистые ленты, несущие пыльцу, приклеиваются к телам насекомых. Эту пыльцу они переносят на рыльца цветков других соцветий. Во время раскрывания пыльников температура початка значительно повышается, у монстеры деликатесной обычно на 15° по сравнению с температурой окружающей среды. Через несколько дней после этого покрывало ее соцветия подвядает и опадает, початок же в длительном ходе развития плодов значительно увеличивается в размерах. Ягоды монстеры кисло-сладкие, пахнущие ананасом. Название "монстера деликатесная" дано этому виду за съедобные плоды. Однако не спешите их попробовать! Даже чуть недозревшие плоды вызывают сильное жжение во рту, из-за обилия игольчатых трихосклереидов и рафид, растворяющихся только при достижении плодом полной зрелости.
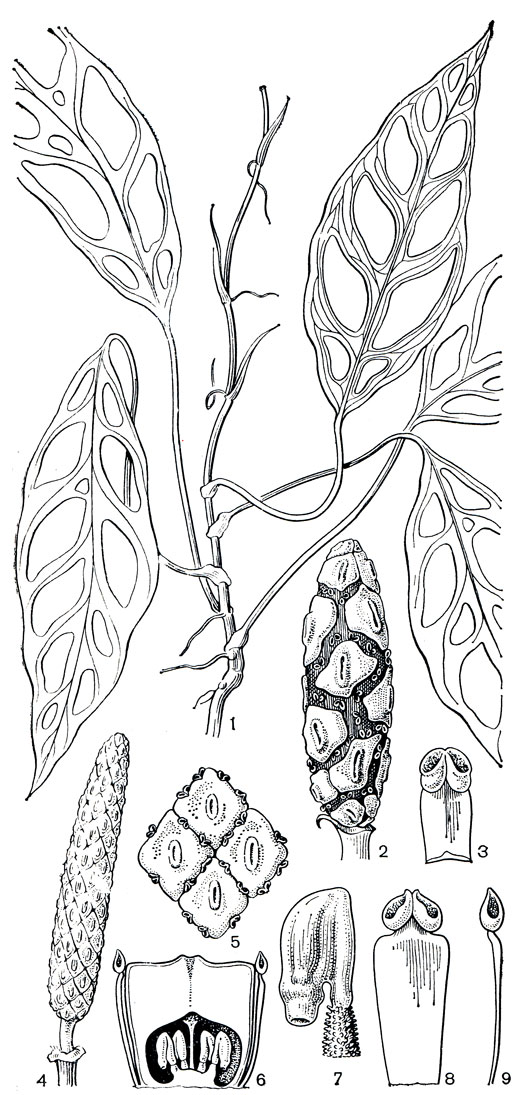
Рис. 267. Аронниковые: подсемейство монстеровые. Монстера неравнобокая (Monstera obliqua): 1 - ветвь с листьями; 2 - початок; 3 - тычинка. Стеносперматион Спруса (Stenospermation spruceanum): 4 - початок (женская стадия цветения); 5 - четыре цветка в мужской стадии цветения; 6 - продольный разрез цветка; 7 - семязачаток; тычинка: 8 - вид спереди; 9 - вид сбоку
Монстера деликатесная - декоративное растение, у нас широко культивируется и привлекает внимание не только крупными продырявленными листьями, но и придаточными воздушными корнями, во множестве развивающимися преимущественно на горизонтальных участках стебля. Корни свободно свисают, иногда образуя подобие живого занавеса, их покровная ткань (веламен) всасывает конденсирующуюся влагу. Эта монстера так широко распространена в культуре, что теперь уже трудно установить, где ее родина. Другие широко культивируемые виды монстер (монстера Адансона - М. adansonii, монстера сомнительная - М. dubia) имеют четко очерченные естественные ареалы, и в их пределах местное население использует эти растения как лекарственные.
Калловые, или белокрыльниковые (Calloideae),- самое северное и единственное подсемейство аронниковых, не имеющее своих представителей в тропиках. Это небольшая группа из 4 родов, включающая всего 5 видов, распространена в умеренной зоне северного полушария, вплоть до субарктических районов. Калловые - корневищные травы с простыми цельнокрайными листьями и обоеполыми с околоцветником цветками (голые только у белокрыльника - Calla). В отличие от уже рассмотренных групп калловые имеют членистые млечники. Наиболее широко распространен и хорошо известен монотипный род калла, или белокрыльник. Толстые водянистые стебли его единственного вида - белокрыльника болотного (С. palustris) - несут небольшие сердцевидные листья. От длинного корневища отходят мощные втягивающие корни, удерживающие растение в зыбком субстрате заболоченных почв. На неярком фоне болот и заболоченных лугов он весной привлекает внимание белизной покрывала соцветия, позднее - интенсивно красными плодами, собранными в густые короткие соплодия (рис. 268, табл. 63). Плоды распространяются преимущественно водоплавающими птицами. Семена белокрыльника очень мелкие, их оболочка имеет хорошо развитую воздухоносную ткань с заполненными воздухом межклетниками; благодаря этой особенности они легко разносятся водой и не теряют плавучести в течение многих месяцев.
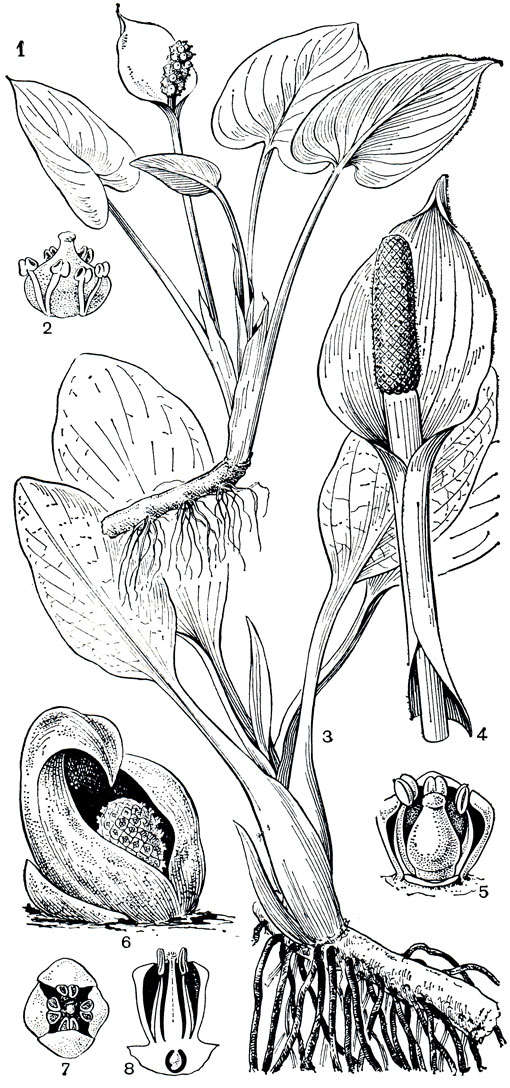
Рис. 268. Аронниковые: подсемейство калловые. Белокрыльник болотный (Calla palustris): 1 - общий вид; 2 - цветок. Лизихитон камчатский (Lysichiton camtschatcense): 3 - общий вид растения; 4 - соцветие; 5 - цветок. Симплокарпус вонючий (Symplocarpus foetidus): 6 - соцветие; 7 - цветок (вид сверху); 8 - продольный разрез цветка
Два близких вида рода лизихитон (Lysichiton) представлены мощными растениями с крупными (до 2 м) листьями и снежно-белым (лизихитон камчатский - L. camtschatcense) или кремовым (лизихитон американский - L. аmеricanum) покрывалом. Оба вида растут на болотах, заболоченных лугах и интенсивно разрастаются на вырубках березовых лесов.
К этому же кругу родства относится симплокарпус (Symplocarpus) - монотипный род, распространенный в Восточной Азии и на востоке Северной Америки. Для наших северных широт симплокарпус не совсем обычное растение. Ранней весной его яркие соцветия первыми появляются на буром фоне прошлогодней травы. Малиново-красные или лиловые мясистые покрывала имеют конусовидную форму и на-поминают шлемы (основание покрывала и цветонос погружены в почву). Оттенки и интенсивность окраски покрывала варьируют в зависимости от степени содержания разных антоциановых пигментов. Внутри такого шлема-покрывала виден почти округлый желтоватый початок с обоеполыми цветками (табл. 62). Симплокарпус привлекает насекомых ярким цветом покрывала и чесночным запахом. Его посещают многочисленные мухи, главным образом горбатки из рода фора (Phora), пчелы, небольшие клопы, не исключена возможность опыления улитками и самоопыления. Цветет симплокарлус в безлистном состоянии и только после отцветания, одновременно с формированием плодов, начинают быстро развиваться его ярко-зеленые широкосердцевидные или широко-копьевидные листья. Растет симплокарпус на лугах, по берегам рек и ручьев, на заболоченных местах, нередко прямо в воде. На юге советского Приморья он известен также на заболоченных приморских лугах, где несколько угнетен густым травяным покровом. Значительно лучшего развития он достигает там в редкостойных ольшаниках из японской ольхи (Alnus japonica). В травяном покрове этих лесов господствуют гигантские розетки симплокарпуса, образованные листьями почти метровой длины. Округлое соцветие-початок к концу лета превращается в крупное шаровидное соплодие диаметром 10-20 см и массой до 300 г. На поверхности соплодия как бы вырезан четкий геометрически правильный узор, образованный затвердевшими частями околоцветника и рылец; семена глубоко погружены в разросшуюся мясистую ось початка. Под тяжестью разрастающейся оси соцветия и развивающихся плодов плодонос початка сгибается и размещенное в основании растения соплодие опускается прямо на почву. Плоды части соплодия, непосредственно контактирующие с влажной почвой, в большинстве случаев развиваются медленней или вообще остаются недоразвитыми. В результате соплодие теряет шаровидную форму, становится асимметричным, неправильно продолговатой или даже сердцевидной формы.

Таблица 62. Аронниковые: 1 - алоказия пахучая (Alocasia odora), оранжереи Ботанического института АН СССР в Ленинграде; 2 - стеносперматион попаянский (Stenospermation popayanense), там же; 3 - аглаонема изменчивая (Aglaonema commutatum), там же; 4 - симплокарпус вонючий (Symplocarpus foetidus), общий вид растения, парк Ботанического института АН СССР в Ленинграде; 5 - соцветие симплокарпуса вонючего, там же
В Северной Америке симплокарпус называют восточной скунсовой капустой, противопоставляя ему лизихитон, как западную скунсовую капусту. Изучавший эти растения американский ботаник Г. Турессон (1916) сообщает, что оба растения распространяют зловоние, которое можно сравнить лишь с тошнотворным запахом жидкости (секрета) скунса. Судя по литературным данным, распространенный у нас лизихитон камчатский вообще не имеет запаха, а неприятный запах, свойственный симплокарлусу, очень слаб и уж никак не вызывает "скунсовых ассоциаций". Перезрелые плоды симплокарпуса издают приятный аромат, очень похожий на запах соплодий джекфрута (Artocarpus heterophyllus) - ближайшего родича хлебного дерева. Отдаленно этот запах напоминает аромат ананаса. Мякоть соплодий симплокарпуса не только ароматична, но и имеет сладкий вкус. Все это не оставляет сомнений, что симплокарпус принадлежит к зоохорным растениям. В Северной Америке его семена были найдены в желудках диких уток. Симплокарпус нередко считают ядовитым растением, засоряющим пастбища. Скот его не ест, и лишь в Америке им безнаказанно питаются древесные крысы. Симплокарпус широко применялся в народной медицине как антисептик. Американские индейцы использовали корневые волоски его для лечения зубной боли, корни - для татуировки и как талисман, против возвращения болезней. Используют симплокарпус и как пищевое растение, но только вареным (после отваривания неприятный запах растения исчезает), из высушенных корней приготавливают муку для выпечки хлеба, отваренные молодые листья заменяют овощи.
Последний представитель подсемейства - оронтиум водный (Orontium aquaticum) - единственный вид наиболее обособленного и специализированного рода среди калловых.
Оронтиум - небольшое травянистое растение с глубоко сидящим вертикальным корневищем и продолговато-эллиптическими, обычно плавающими листьями. Обитает он в заводях, ручьях, трясинах и болотах, чаще всего развиваясь как типичное водное растение. Над водой возвышаются его золотистые булавовидные початки, благодаря им это растение называют в Америке "золотая дубинка". Развитие плодов сопровождается постепенным отклонением от растения-плодоноса, в результате чего формирующееся соплодие погружается в воду. Как только плоды созревают, они отрываются от початка и сразу же всплывают; плавучесть их поддерживает толстый околоплодник. Примерно через неделю (этого времени бывает достаточно, чтобы плоды оказались далеко от материнского растения) околоплодник наполняется водой и плод опускается на дно. Еще через такой же промежуток времени околоплодник разлагается и семя прорастает. Местное население использует оронтиум как пищевое растение: едят его обжаренные семена и отваренные корневища.
Ареал оронтиума сравнительно небольшой, но тоже дизъюнктивный. Он распространен в Северной Америке на побережье Атлантической низменности, а также в удаленных от моря районах Аппалачей, где заходит в горы до высот 800-840 м над уровнем моря.
В подсемействе лазиевые (Lasioideae) около 15 родов и более 150 видов тропических растений Старого и Нового Света. Это преимущественно гигрофильные многолетние травы, нередко исполинские, с корневищами или клубнями, их листья от цельных стреловидных до 3-раздельных с перисторассеченными сегментами. У представителей многих родов растение имеет лишь один, часто гигантский лист; на черешках листьев и на цветоносе обычны многочисленные схизогенные вместилища, особенно у развивающихся в водной среде видов из родов лазия (Lasia), циртосперма (Cyrtosperma), уроспата (Urospatha). Цветки 2-3 - членные, протогиничные, обоеполые обычно с околоцветником, однополые - голые. Тычинок 4-6; гинецей из 2-5 плодолистиков или псевдомономерный, завязь 1-гнездная, с 1-2, реже несколькими семязачатками. У представителей пантропического рода циртосперма и палеотропического подолазия (Podolasia) пыльники раскрываются во всех цветках соцветия почти одновременно и у многих цветков обычно самоопыление.
В подсемействе по крайней мере 2 рода представлены лазающими растениями: это эндемики Гвинео-Конголезской флористической области роды церцестис (Cercestis) и ректофиллум (Rhektophyllum). Распространенный в прибрежных районах тропической Западной Африки ректофиллум удивительный (R. mirabile) на ювенильных побегах развивает небольшие стреловидные листья, на лазающих побегах - крупные рассеченные лопастные листья.
Лазиевые обычно растут отдельными, изолированными экземплярами и не образуют зарослей. Монтричардия древовидная (Моntrichardia arborescens) является в этом отношении исключением. Это растение развивает прямой, похожий на ствол дерева стебель, достигающий в высоту 3 м, увенчанный крупными стреловидными листьями с примитивным жилкованием. У ее колючей разновидности (М. arborescens var. aculeata) стебель покрыт крупными шипами и достигает в высоту 7-9 м. Монтричардия - пионерное растение. Известный исследователь тропических лесов П. Ричарде (1961) считает его первым поселенцем на илистых отложениях проток дельтовой зоны Амазонки. Плоды монтричардии поедают и разносят главным образом птицы, обитающие в заливах и болотах Южной Америки. Попавшие на подходящий субстрат, семена монтричардии прорастают необычайно быстро, и проростки ее могут укорениться в промежутке времени между двумя приливами. Благодаря интенсивному развитию всходов растение за короткий срок образует почти чистые заросли, чередующиеся с зарослями из дрепанокарпуса луновидного (Drepanocarpus lunatus) из семейства бобовых. Под прикрытием этих зарослей начинают развиваться мангры, впоследствии заглушающие монтричардию, и она переходит в положение угнетенного подлеска. Стадия зарослей монтричардии древовидной характерна для зарастания песчаных берегов рек в бассейне Амазонки.
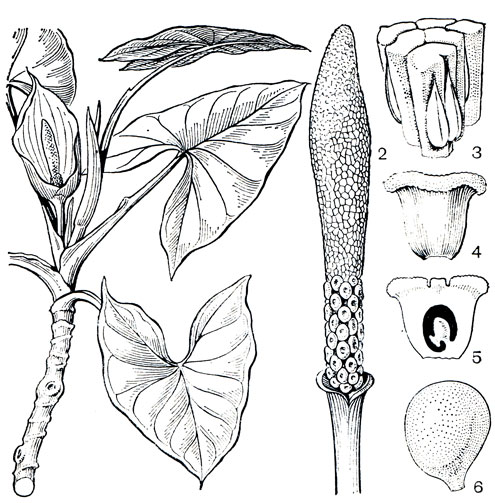
Рис. 270. Монтричардия древовидная (Montrichardia arborescens): 1 - верхняя часть цветущего побега; 2 - початок (внизу - женская часть, вверху - мужская часть соцветия); 3 - мужские цветки (вид сбоку); 4 - женский цветок - гинецей (вид сбоку); 5 - продольный разрез гинецея; 6 - плод
Среди лазиевых, несомненно, наиболее интересен самый крупный род подсемейства - аморфофаллус - абориген тропических лесов и саванн Старого Света, особенно травянистый великан аморфофаллус гигантский. Немногим более ста лет назад (в 1878 г.) итальянский ботаник Одорадо Беккери нашел это растение в дождевом тропическом лесу Западной Суматры. Впоследствии его удалось вырастить в оранжереях ботанических садов ряда стран, и это растение-чудо с огромным, значительно превышающим рост человека соцветием произвело сенсацию и вызвало паломничество в ботанические сады. Журналисты, неоднократно писавшие об аморфофаллусе, называли его соцветие "самым крупным цветком в мире". Огромное чашеобразное покрывало, причудливо окрашенное в зеленовато-беловатые и красные тона, гофрировано в верхней части и скрывает нижнюю половину початка, несущую в основании женские цветки (рис. 269). Над ними располагается около 5 тыс. мужских цветков. Из центра покрывала примерно на 1,5 м возвышается в виде мощного конуса самая верхняя, стерильная часть початка. Во время цветения она значительно нагревается, и именно в этот период от нее исходит крайне неприятный запах. У отдельных растений зловоние настолько сильно, что ухаживающие за ними сотрудники, чтобы избавиться от запаха, вынуждены менять одежду. Растение цветет в безлистном состоянии, и только после образования плодов появляется его единственный лист. Черешок такого листа, несколько расширяющийся книзу, больше похож на ствол небольшой пальмы (он может достигать в высоту 2-5 м!), а сложнорассеченная пластинка листа - на ее крону. Каждая из частей 3-раздельной пластинки листа дважды перисторассечена, и длина каждого из этих сегментов может достигать 2 м. Конечно, такие гигантские органы развиваются не сразу из семени, им предшествует целый ряд сменяющих друг друга генераций листьев и клубней. Причем с каждой новой генерацией увеличиваются размер и рассеченность нового развивающегося листа, диаметр и масса клубня. Наконец, когда в клубне накоплено достаточно питательных (запасных) веществ, развивается соцветие. Первое соцветие тоже еще сравнительно небольшое (меньше метра), последующие же генерации соцветий, листьев и клубней становятся все крупнее. По-видимому, близка к предельной высота соцветия в 2,6 м, известен 5-метровый черешок листа и клубень массой 40 кг при диаметре примерно 50 см.
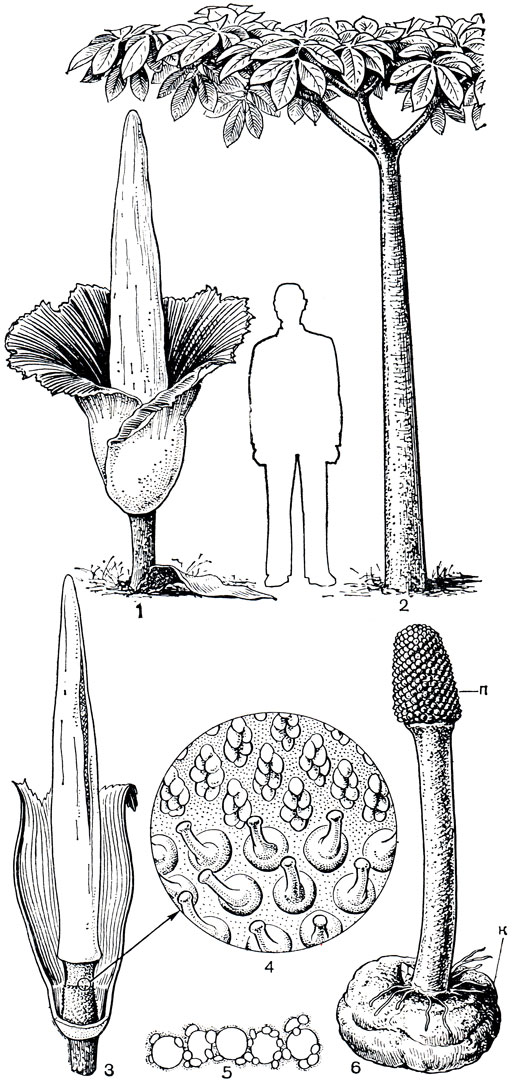
Рис. 269. Аморфофаллус гигантский (Amorphophallus titanum). Общий вид растения: 1 - в стадии цветения (соцветие); 2 - в вегетативной стадии (единственный лист); 3 - соцветие (передняя часть покрывала удалена); 4 - фрагмент соцветия в пограничной зоне между женскими (внизу) и мужскими (вверху) цветками; 5 - поллиний; 6 - растение в стадии плодоношения: п - соплодие, к - клубень
Весь процесс цветения занимает у аморфофаллуса несколько дней. Опыление цветков производят попадающие в огромную чашу по-крывала соцветия навозные жуки, привлеченные запахом. Затем, буквально на глазах, чаша покрывала сморщивается, подвядает и отваливается вместе с верхней частью початка. На коротком цветоносе остается лишь зона женских цветков. Завязи цветков постепенно разрастаются, образуя мясистые ягоды, распространяемые животными.
Подсемейство филодендровые (Philodendroideae) включает примерно 15 родов и более 500 видов. Это преимущественно лазающие растения, реже древовидные или травянистые формы. Характерно образование корневищ, придаточных воздушных корней, в тканях обычны млечники, в клетках - рафиды. Цветки однополые, голые, женские иногда со стаминодиями; тычинки часто в синандриях; гинецей из 2-14 плодолистиков; завязь одно- многогнездная, с одним - многочисленными семязачатками. Плод - сочная ягода.
Филодендровые - энтомофильные, зоохорные растения, широко распространенные в тропической зоне, но распределены в пределах ее очень неравномерно. Богаты ими американские и азиатские тропики. В Африканском флористическом подцарстве встречаются лишь небольшие, как правило, обособленные своеобразные роды. Основной род подсемейства филодендрон, включающий более 250 видов, распространен только в Неотропическом царстве. Большинство его видов обитает в нижнем горном поясе и на низменностях и представлено преимущественно лазающими растениями. Переход к лазанию имеет глубокий биологический смысл. В дождевых тропических лесах до поверхности почвы доходит ничтожное количество света, но уже на высоте 2,5 м освещенность почти удваивается, поэтому лазающие растения, если они даже, как аронниковые, не пробиваются в верхний полог, значительно выигрывают в световом режиме по сравнению с растениями нижнего яруса. И это размещение лазающих растений в условиях лучшего освещения осуществляется без построения ими самоподдерживающих структур (мощных стволов и прочее), требующих больших энергетических затрат. В отличие от антуриумов, филодендроны не отличаются большим разнообразием горных форм и не заходят высоко в горы. Высотное распространение их обычно ограничивается поясом 1500-2000 м над уровнем моря, но и на этих высотах произрастает сравнительно немного видов. Жизненные формы филодендронов разнообразны, не менее разнообразны и морфологические признаки этого большого рода. Листья от цельных до дважды перисторассеченных, черешки с влагалищами. Покрывало соцветия толстое, мясистое, с перетяжкой, свернутое в трубку, приоткрывающуюся во время цветения и обычно развертывающуюся после оплодотворения цветков. Початки цилиндрические с очень тесно расположенными цветками. В нижней части размещаются женские цветки, каждый из них представлен гинецеем: яйцевидной 2 - многогнездной завязью
и преимущественно полушаровидными рыльцами. Выше по початку сравнительно небольшая зона стерильных (мужских) цветков и оканчивается початок мужскими цветками с 2-6 необычной формы тычинками. Тычинки сидячие, с сильно разросшимся связником, образующим подобие геометрической фигуры типа призмы или перевернутой усеченной пирамиды. На боковых сторонах этого образования размещены пыльники. Тычинки так тесно расположены на початке, что их границы, как и границы мужских цветков, почти неразличимы. У лазающего филодендрона имбе (P. imbe), широка культивируемого в оранжереях, границы между мужскими цветками проступают в виде тонкой, неясной сетки, едва заметной на белой поверхности початка. Развивающееся соцветие находится внутри зеленоватого покрывала, полностью скрывающего початок. Лишь к началу цветения края покрывала расходятся, его внутренняя поверхность становится красновато-лиловатой и на ее фоне отчетливо выступает снежно-белая, как бы восковая верхняя часть початка, несущая мужские цветки. В это время начинается цветение расположенных в нижней части початка женских цветков. В углублениях их рылец появляются капли ярко-оранжевой вязкой жидкости - это и есть воспринимающая пыльцу поверхность, явно привлекающая внимание насекомых-опылителей. Спустя некоторое время рыльца подсыхают (обычно не все сразу) и начинается мужская фаза цветения. Сжатые и прикрытые разросшимися связниками пыльники начинают разрастаться - раздвигают преграды, в данном случае разъединяют тычинки. Линии их разграничения, до сих пор едва заметные, становятся все более отчетливыми, наконец, между тычинками появляются щели. Примерно в этот момент пыльники раскрываются в верхней части и пыльца под давлением извергается. Но у филодендрона имбе, как и у многих других аронниковых, пыльца склеена в липкие ленты, они-то и выдавливаются из пыльников, как паста из тюбика, образуя на початке массу кудрявых червеобразных нитей (рис. 271). Вскоре некоторые нити отрываются, попадают на нижерасположенные женские цветки и опыляют их, если рыльца еще не потеряли восприимчивость. Но большинство нитей приклеивается к телам ползающих по початку насекомых-опылителей, они и переносят пыльцу на цветки других растений.
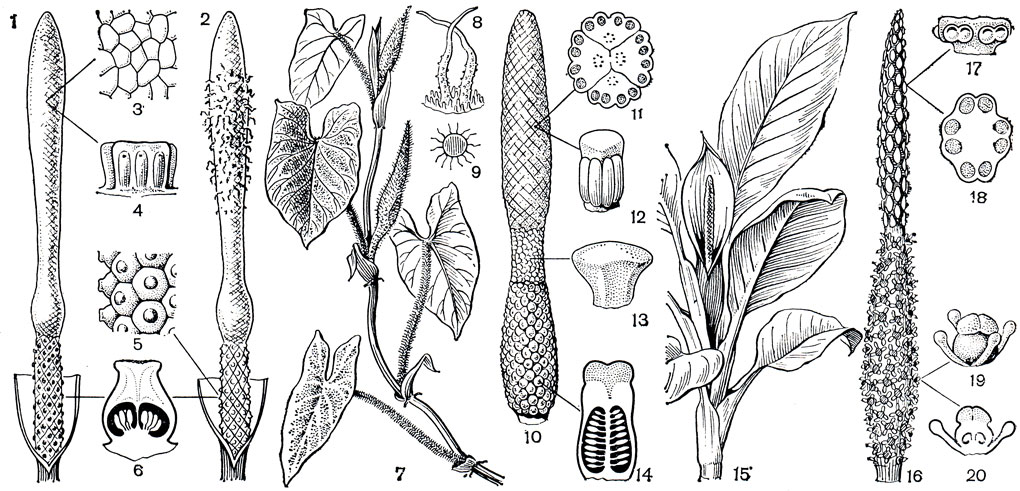
Рис. 271. Аронниковые: подсемейство филодендровые. Филодендрон имбе (Philodendron imbe): 1 - початок в женской стадии цветения (покрывало удалено); 2 - початок в мужской стадии цветения; часть соцветия с мужскими цветками: 3 - вид сверху; 4 - вид сбоку; 5 - часть соцветия с женскими цветками; 6 - продольный разрез гинецея (объяснения в тексте). Филодендрон бородавчатый (P. verrucosum): 7 - общий вид; 8 - волоски на черешках листьев; 9 - поперечный разрез черешка листа; 10 - початок; 11 - поперечный разрез мужского цветка; 12 - тычинка; 13 - стерильный цветок; 14 - продольный разрез завязи. Диффенбахия приземистая (Diffenbachia humilis): 15 - общий вид; 16 - початок; 17 - мужской цветок (вид сбоку), тычинки соединены в синандрии; 18 - то же на поперечном срезе; 19 - женский цветок с 4 стаминодиями; 20 - то же в продольном разрезе
На родине, в Южной Бразилии, филодендрон имбе используют главным образом в народной медицине как антисептик. Его воздушные корни используют как веревки, из коры плетут корзины.
Филодендрон дваждыперистонадрезный (Р. bipinnatifidum) - оригинальное древовидное растение с прямостоячим коротким узорчатым стволом, его геометрически правильный рисунок образуют светлые овалы - следы от опавших листьев. Стебель оканчивается крупными дваждыперистыми листьями на длинных черешках. Интересные наблюдения над его цветением в естественных условиях обитания провел известный датский ботаник И. Э. Варминг (1883). Он подметил несколько любопытных явлений, протекающих последовательно или почти одновременно в период его цветения. Раскрывание покрывала соцветия сопровождается повышением температуры пыльников на 12,5° по сравнению с температурой окружающей среды. Этому сопутствует выделение соцветием очень сильного, одуряющего запаха, привлекающего множество насекомых-опылителей, главным образом мелких жуков, которые проникают в соцветие и остаются там некоторое время, вероятно привлеченные ароматической, вязкой, желтоватой жидкостью, выделяемой в этот момент секреторными клетками на внутренней поверхности покрывала в области женского соцветия. Ползая в основании соцветия, насекомые могут опылять цветки пыльцой, принесенной на их тельцах. Во время формирования плодов верхняя часть соцветия, как и у других аронниковых, подвядает и опадает, нижняя, несущая завязи, сильно разрастается и образует плотное соплодие из желтых 5-6-семянных ягод, увенчанных черноватыми рыльцами. Кисло-сладкие плоды распространяют животные, особенно часто обезьяны и летучие мыши. Этот декоративный вид, культивируемый у нас в оранжереях, широко распространен в лесах Южной Бразилии на высоте 200-300 м над уровнем моря. Местное население использует его в различных целях: жгучий млечный сок как лекарственное средство, ягоды - излюбленное лакомство, воздушные корни идут на приготовление грубых веревок, деревянных гвоздей.
Декоративно и своеобразно небольшое лазающее растение - филодендрон бородавчатый (P. verrucosnm), привлекающий внимание красными черешками, несущими длинные причудливо изогнутые чешуйчатые волоски (рис. 271, 8), стреловидными листьями, сверху зелеными, снизу с красновато-лиловатыми полосами, и красноватыми соцветиями. Естественно распространен этот вид в Латинской Америке на высоте до 1000 м над уровнем моря. С этими же районами связано произрастание другого декоративного лазающего вида - филодендрона краснеющего (P. erubescens). Он имеет темно- пурпурные соцветия (табл. 63) и красноватые междоузлия.
Среди представителей рода диффенбахия (Diffenbachia), распространенных по всей тропической Америке, много ядовитых растений, к ним относится и полиморфный вид диффенбахия сегуина (D. seguina). Жгучий сок, содержащийся в растении, вызывает сильные ожоги слизистых оболочек и кожи. В Вест-Индии в прошлом этим растением плантаторы наказывали рабов, заставляя их откусывать куски стебля. Возникающая сразу же опухоль слизистых оболочек и языка затрудняет речь, откуда и народное название растения, в переводе означающее "немая розга". Природа токсичности этого растения, как указывает американский ботаник Т. Плоуман (1969), еще полностью не раскрыта. Предполагают, что это - результат сов-местного механического и химического воздействия. При откусывании растения миллионы рафид проникают в слизистую оболочку и ранят ее, химическая же токсичность приписывается необычайно ядовитой протеиновой фракции, быстро проникающей в образовавшиеся ранки. Кроме того, во всех частях растения содержатся цианистые соединения. Растение использовалось местным населением как продукт питания, целебное средство, как яд для истребления грызунов и насекомых. Пытались найти применение этому растению и в период второй мировой войны и отнюдь не в гуманных целях. Т. Плоуман пишет, что один из лидеров нацизма - Гиммлер, получив сведения о возможности использовать экстракт из "немой розги" для стерилизации узников концентрационных лагерей, поручил собрать всю информацию об этом растении и начать разведение его в специальных оранжереях непосредственно в концентрационных лагерях. Однако вскоре Бразилия объявила войну фашистской Германии и получить "немую розгу" из Америки оказалось невозможным. План широкого использования этого растения в изуверских целях не реализовался.
Несколько обособленное место в подсемействе занимает монотипный островной род тифонодорум (Typhonodornm), распространенный в Мадагаскарском флористическом подцарстве и на острове Занзибар. Тифонодорум Линдли (Т. lindleyanum) - крупное травянистое растение (высотой до 4 м), образующее густые заросли в болотах, реках и ручьях (рис. 272). Нижняя половина его стебля погружена в воду, надводная часть оканчивается пучком крупных листьев со стреловидной пластинкой длиной более метра и мощными черешками с широкими влагалищами. На длинном (до 50 см) початке размещены 4 зоны цветков. Женские цветки, состоящие из гинецеев, окруженных стаминодиями, расположены в нижней зоне; выше по початку - короткая зона стерильных цветков, представленная синандродиями (сросшимися стаминодиями); ее сменяют мужские цветки каждый из 4-8 тычинок, и оканчивается початок зоной стерильных цветков из 3- 6 стаминодиев каждый. Ценокарпный гинецей образован 4-5 плодолистиками, что отчетливо видно по еще несросшимся 4-5 рыльцам, но завязь уже одногнездная, с 1-2 ортотропными семязачатками (рис. 272, 4). Совершенно своеобразно развиваются семена тифонодорума. Здесь встречается особый тип живорождения, существенно отличающийся от типичной вивипарии, с которой читатель уже познакомился на примере мангров (Жизнь растений, т. 5, ч. 2, с. 232). Зародыш тифонодорума образует листовые зачатки, находясь еще внутри плода и семенной кожуры (рис. 272, 7). Почечка зародыша получает возможность развиваться в семени за счет питательных веществ эндосперма, которые поставляет ей с помощью гаустории семядоля. Находясь внутри семенной кожуры, почечка успевает сформировать 3-4 узких зачатка листа и придаточные корни. Когда плод созреет, трубка покрывала отрывается от растения и падает в воду; в воде оказываются и семена растения, долгое время сохраняющие плавучесть. После укоренения у зародыша развиваются сначала такие же узкие листья и лишь постепенно формируются стреловидные, характерные для взрослого растения.
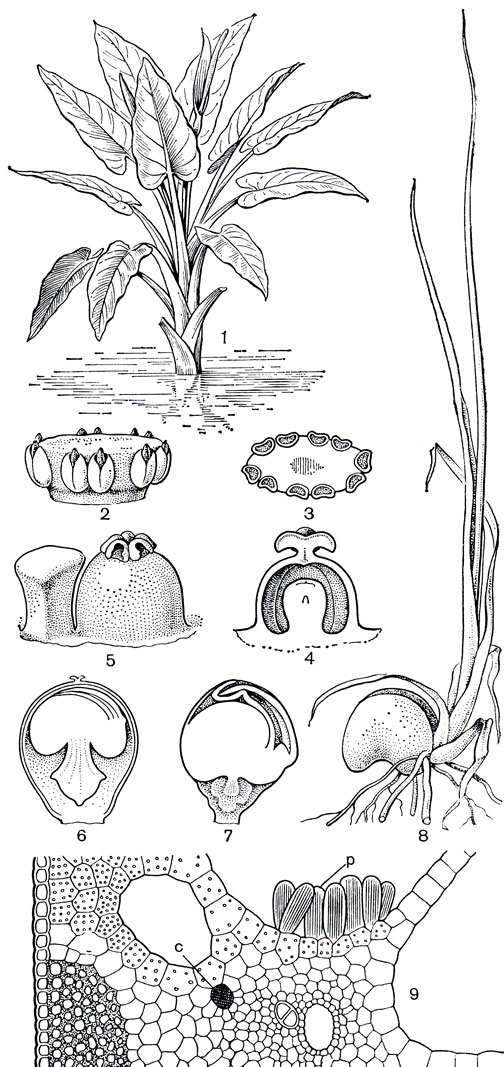
Рис. 272. Тифонодорум Линдли (Typhonodorum lindleyanum): 1 - общий вид; 2 - мужской цветок (синандрий) в стадии начала раскрывания пыльников; 3 - то же на поперечном срезе; 4 - продольный разрез гинецея; 5 - гинецей и стаминодий; 6 - продольный разрез плода; 7 - развивающиеся в семени зародыш с гаусторией, внедрившейся в эндосперм, и зачатками листьев; 8 - проросток; 9 - поперечный срез нижней части жилки листа; р - группа клеток с рафидами, выступающими в межклетники, с - секреторная клетка
Тропическое подсемейство колоказиевых (Colocasioideae) включает множество ценных пищевых растений, имеющих огромное значение в питании населения тропических и субтропических стран. Примерно 15 родов и более 150 видов объединяет эта группа крупных трав с хорошо выраженными подземными стеблями - мощными клубнями, толстыми корневищами. У ряда видов развиваются толстые, короткие надземные стебли, обычно не ветвящиеся, но чаще их нет и наземные органы представлены крупными простыми, цельнокрайными листьями на длинных черешках и цветоносными побегами. Антоцианы, содержащиеся в листьях, придают им красновато-лиловый оттенок (особенно у молодых листьев), вызывают пестролистность, иногда с причудливым распределением красок. В стеблях, листьях, клубнях обычны млечники, много клеток с рафидами. Толстые плотные покрывала соцветий имеют характерную перетяжку, ниже ее на початке расположены женские цветки с одногнездной завязью и несколькими семязачатками, выше - мужские, состоящие из сросшихся в синандрии тычинок; пыльники раскрываются порами, пыльцевые зерна объединены в червеобразные слизистые ленты. В промежуточной зоне, под перетяжкой покрывала соцветия на початке формируются стерильные цветки со сросшимися в синандродии стаминодиями. У большинства колоказиевых покрывало в самой узкой части, в зоне перетяжки, почти примыкает к початку и в оставшуюся узкую щель могут проникнуть к женским цветкам только очень мелкие насекомые. Затруднения, встречаемые опылителями на этом пути, возможно, одна из причин редкого образования полноценных плодов. Вегетативное размножение у колоказиевых явно преобладает, если не становится единственным у ряда видов. Особенно редко плодоносят широко культивируемые виды, разводимые обычно подземными стеблями.
Первое место среди используемых видов, несомненно, принадлежит таро - так называют клубни и само растение колоказии древней (Соlocasia antiqnornm), известной также под названием колоказии съедобной (С. esculenta). Это травянистое растение развивает крупные сердцевидные листья с черешками длиной до 2 м и мощные крахмалистые клубни, ради которых его и разводят уже более 2000 лет. Есть много предположений о родине таро, и, по-видимому, Южная Азия наиболее вероятное место его происхождения. В Китай таро проникло в I в. н. э. и распространилось по островам Тихого океана. На Гавайях и Таити его разведение было особенно успешным, множество культурных форм этого растения существует там и сейчас. Утверждают, что само название "таро" - таитянская форма яванского названия растения. Распространялась культура таро и в западном направлении. Из рукописей Плиния известно, что уже в его времена таро использовали в пищу египтяне. Позже оно появилось в тропической Западной Африке, откуда африканские рабы вывезли его на острова Вест-Индии, а затем таро проникло и на Американский континент. В настоящее время его разводят в тропической и субтропической зонах и в естественном состоянии это растение уже неизвестно.
Имеется множество разновидностей таро, различающихся по форме и окраске листьев, строению соцветий. Но особенно разнообразны его клубни - основной объект деятельности селекционеров. Выведено множество сортов таро, различающихся по форме и строению клубней, их размеру и количеству, содержанию в них тех или иных химических веществ, вкусовым качествам и так далее. Обычен у таро один крупный корневищеподобный клубень, но есть сорта и со множеством мелких, окружающих его клубней (рис. 273). Распространение у таро по крайней мере 4 хромосомных рас с числом хромосом в соматических клетках 28, 42, 36 и 48 говорит о разных основных хромосомных числах и о генетической неоднородности растений, объединенных под этим названием.
Таро культивируют на низменностях, на переувлажненных или заболоченных почвах, разводят его и в горах (в Гималаях, например, культура таро встречается и выше 2500 м над уровнем моря). Используют клубни таро примерно в тех же целях и так же разнообразно, как картофель в умеренных широтах. Таро всегда едят в вареном или жареном виде, сырые клубни его вызывают ощущение сильного жжения во рту. Одной из причин этого считают действие содержащихся в них рафидов. Рафиды расположены в плотных пучках в специальных цилиндрических клетках. Если конец такой клетки разрушается, игольчатые кристаллы под давлением клеточного сока с силой извергаются и, по-видимому, ранят слизистую оболочку рта. Кроме клубней, у колоказии древней используют в пищу также толстые, мясистые корневища и листья.
В тропиках Нового Света естественно распространено более 40 видов рода ксантосома (Xanthosoma). Заслуживает внимания вест-индская ксантосома стрелолистная (X. sagittifolium), чаще всего называемая там "маланга". Растение это разводили еще в доколумбовой Америке. Культурный ареал этого вида в настоящее время охватывает тропики обоих полушарий. Во взрослом состоянии ксантосома имеет толстый прямой наземный стебель более метра высоты и превышающие его по длине стреловидные листья с черешками до 2 м. Покрывало соцветия, зеленое снаружи и кремовое внутри, в верхней части открыто, в нижней свернуто в трубку, внутри которой находится небольшой початок (рис. 273). В его основании расположены женские цветки. Их гинецей образован 3-4 плодолистиками, рыльце дисковидное, завязь 2-4-гнездная, с многочисленными семязачатками. Выше по початку - зона стерильных цветков, образованных синандродиями, и оканчивается початок мужскими цветками, их тычинки по 4-6 срастаются в синандрий. Разводят ксантосому главным образом из-за толстых клубневидных корневищ, но в пищу используют также и надземный стебель и листья. Экономическое значение как источники питания имеют также ксантосома Жакэна (X. jacquinii, рис. 260), ксантосома фиолетовая (X. violaceum), ксантосома мощная (X. robustum) и др. Среди видов, распространенных в Палеотропическом царстве, заслуживает внимания самый крупный в подсемействе (более 60 видов) род алоказия (Аlоcasia), среди представителей которого немало ядовитых растений. Использование их в пищу требует специальной предварительной обработки. Наиболее широко используется в различнейших целях естественно произрастающая в Юго-Восточной Азии алоказия крупнокорневищная (A. macrorrhiza). Ее разводят как декоративное, пищевое и лекарственное растение в азиатских тропиках вплоть до Новой Каледонии, а также в тропической зоне Нового Света. Однако и у нее некоторые разновидности содержат почти во всех частях растения цианистые соединения.
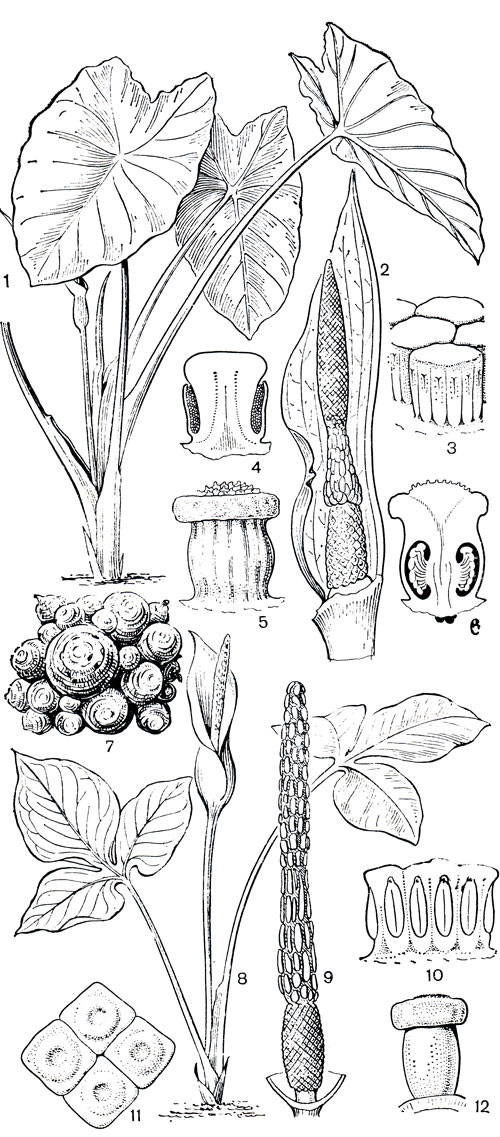
Рис. 273. Аронниковые: подсемейство колоказиевые. Ксантосома стрелолистная (Xanthosoma sagittifolium): 1 - общий вид; 2 - соцветие (часть покрывала удалена); 3 - мужские цветки, состоящие из соединенных в синандрии тычинок; 4 - продольный разрез синандрия; 5 - гинецей; 6 - продольный разрез завязи. Колоказия древняя (Colocasia antiquorum), или таро: 7 - клубни широко культивируемой формы 'дашин'. Ксантосома широк опастная (Xanthosoma platylobum): 8 - общий вид; 9 - початок (покрывало удалено); 10 - синандрии (вид сбоку); 11 - 4 женских цветка (вид сверху); 12 - гинецей
Своеобразную группу живородящих растений (или ложноживородящих, поскольку рассматриваемое явление не связано с прорастанием семян на растении) образуют роды ремузатия (Remusatia) и гонатантус (Gonathantus). Это небольшие растения, высотой обычно до 50 см, иногда развивающиеся как эпифиты на деревьях. Их сердцевидные на длинных черешках листья отходят от слабо сплюснутых клубней. Покрывало соцветия скрывает початок, в верхней части которого размещены синандрии, отделенные зоной стерильных цветков от расположенных в основании початка женских цветков, состоящих только из одного гинецея; завязь одногнездная, содержащая несколько семязачатков. Растения эти плодоносят редко и, по-видимому, в настоящее время размножаются преимущественно вегетативно, для чего и развивают специальные органы. У представителей обоих родов от верхней части клубня отходят ветвящиеся побеги типа столонов с более или менее густо размещенными на них выводковыми почками. Основываясь на первоначально ортотропном положении этих столонов, Карл Гебель (1932) считает их видоизмененными цветоносными побегами. У ремузатии живородящей (R. vivipara) эти побеги красные, выводковые почки на них размещены пучками. Каждая из кроющих почку чешуек крючковидно загнута и этими крючками цепляется за шкуру животного, подобно эпизоохорным плодам и семенам. У ремузатии Хукера чешуйки почки более тонкие, не загнутые, почечки легко отрываются от побега и, по-видимому, распространяются ветром. Достигнув поверхности почвы, почка укореняется и развивается в новое растение. Рассмотренная группа растений в основном жители Гималаев, распространенные на высоте 1000-2000 м над уровнем моря. Ареал ремузатии живородящей более обширен и простирается от тропической Западной Африки, где она распространена на высоте 200-300 м, до Южного Китая.
Среди колоказиевых есть и стелющиеся растения, одно из них - сингониум ножколистный (Syngonium podophyllum). Его побеги стелются по земле, интенсивно ветвятся и укореняются в каждом узле (рис. 260). На ювенильных побегах развиваются характерные стреловидные листья, с возрастом растения сменяющиеся на 5-7-рассеченные. В тканях развиваются млечники, и млечный сок имеется во всех частях растения. Мясистое соплодие сингониума подофиллового в некоторых районах Латинской Америки употребляют в пищу.
Подсемейство аронниковые (Aroideae) - одно из самых специализированных и наиболее интересных среди рассматриваемой группы, распространено преимущественно в тропической и субтропической зонах и включает примерно 30 родов и около 200 видов. Его представители - небольшие прямостоячие наземные травы с клубнями и корневищами, корневищные болотные и водные растения. Их цветки однополые, голые, однодомные и очень редко двудомные; тычинки свободные или в синандриях; завязь преимущественно одногнездная, с несколькими семязачатками; семя с эндоспермом; плод - сочная ягода. Соцветия устроены сложно, их структура включает приспособления для предотвращения самоопыления и привлечения насекомых-опылителей. Характерны сапромиофилия и зоохория. В тканях есть млечные сосуды, в клетках - нередки рафиды, найдены алкалоиды, в том числе и ядовитые. Распространено вегетативное размножение клубнями и корневищами. В нашей стране естественно обитают виды из родов аронник (Arum), аризема (Arisaema) и эминиум (Eminium), привлекающие внимание не только своеобразным внешним обликом, но и необычайно сложными приспособлениями к опылению.
Детально опыление изучено у западноевропейского аронника пятнистого (Arum maculatum) - обычного растения влажных широколиственных лесов, обитающего на богатых гумусом переувлажненных почвах. Строение сложного соцветия аронников можно рассмотреть на рисунке 274. Аронник - сапромиофильное растение, и его опылители мясные, падальные и навозные мухи, как известно, не имеют особых приспособлений для опыления цветков. Поэтому растение "рассчитывает лишь на себя". У него имеется ряд сложнейших приспособлений для привлечения опылителей и достижения эффекта опыления. Характерно, что все эти приспособления сводятся к обману насекомого-опылителя. Растение успешно имитирует цвет субстрата, в который откладывают яйца его опылители. Темные листья и грязно-лилово-красноватое, иногда пятнистое покрывало напоминает разлагающееся мясо. Впечатление усиливают отвратительный запах, исходящий от верхней части (придатка) початка, и его повышенная температура. Обманутые приманкой мухи и мошки из семейства бабочниц (Psychodidae) летят к соцветию. В надежде отложить яйца в подходящий субстрат они вползают внутрь соцветия и оказываются в ловушке. Насекомые не могут удержаться в верхней части соцветия на внутренней поверхности покрывала, несущей скользкие выросты-сосочки, и соскальзывают вниз к основанию соцветия. Ползти вверх им препятствует скользкая поверхность покрывала, к тому же выход из со-цветия закрыт направленными вниз ветвистыми волосками (видоизмененными стерильными цветками). Вынужденные находиться в основании соцветия, насекомые ползают среди женских цветков и поедают выделяющуюся на рыльцевых волосках сладковатую жидкость.
Попутно они опыляют цветки, если перед посещением данного соцветия их тельца уже были осыпаны пыльцой. Но насекомые лишь временные пленники соцветия, и растение совсем небескорыстно их подкармливает. Проходит некоторое время (примерно сутки), и рыльца подсыхают, внутренние стенки покрывала теряют тургор, и находящиеся на их поверхности скользкие сосочки уже не препятствуют насекомым карабкаться вверх. На своем пути к выходу насекомые проползают через зону пылящих тычинок, поскольку к этому времени растение уже вступает в мужскую фазу цветения, пыльники раскрываются, и на тельца насекомых попадает пыльца. Осыпанные пыльцой, насекомые беспрепятственно выбираются наружу: стерильные цветки, ранее закрывавшие выход из соцветия, теперь уже подвяли и не служат помехой. Выйдя из соцветия, насекомые опять дают себя обмануть, и все начинается сначала: они летят к следующему ароннику, попадают в его соцветие и опыляют цветки. Этот способ привлечения опылителей очень эффективен, в ловушку-соцветие попадают сотни и даже тысячи насекомых. Немецкий ботаник П. Кнут (1898) насчитал в одном соцветии аронника пятнистого около 4 тыс. экземпляров бабочницы обычной (Psychoda phalaenoides).
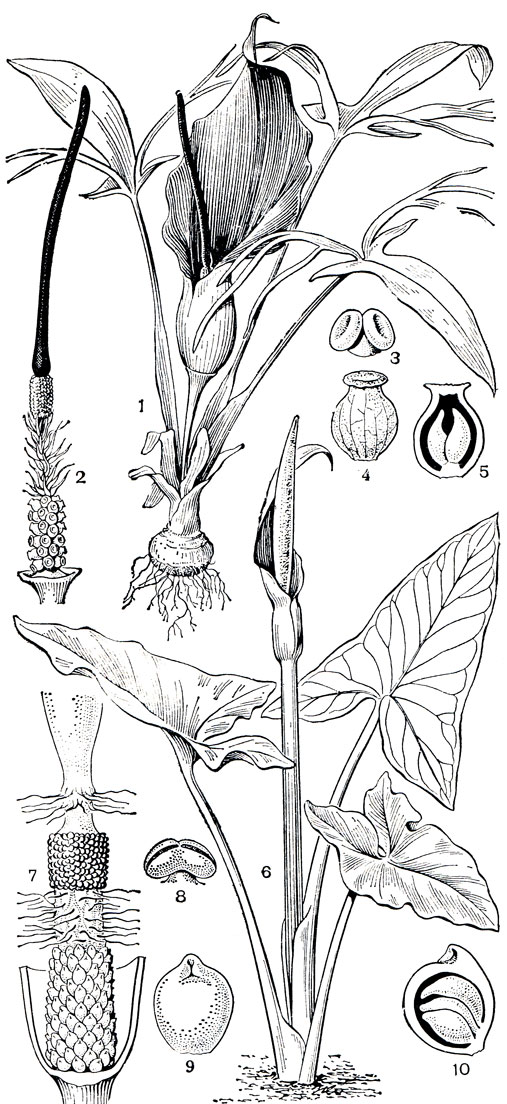
Рис. 274. Аронниковые Средней Азии. Эминиум Альберта (Eminium alberti): 1 - общий вид; 2 - початок (покрывало удалено); 3 - мужской цветок; 4 - женский цветок (гинецей); 5 - продольный разрез гинецея. Аронник Королькова (Arum korolkovii): 6 - общий вид; 7 - соцветие (покрывало удалено); 8 - мужской цветок (тычинка); 9 - женский цветок (гинецей); 10 - плод
Сходно с рассмотренным происходит опыление у интересного растения нашей флоры - аронника восточного (Arnm orientale). Немногим отличается опыление цветков у аронника черного (A. nigrum), обитающего в основном в районах Адриатики. В соцветие-ловушку этого вида попадают многочисленные мухи и жуки. Совершенно другую группу насекомых привлекает аронник конофаллоидный (А. соnophalloides). По указанию К. Фэгри и Л. ван дер Пейла (1966), цветки этого растения опыляют только кровососущие москиты и только самки (самцы не сосут кровь). Предполагают, что соцветие этого аронника привлекает насекомых имитацией запаха шкуры млекопитающего, кровью которого они питаются.
В тропиках и субтропиках Старого Света, от тропической Африки до Филиппин, распространен олиготипный род сауроматум (Sauromatum). Его представители - небольшие травы с единственным пальчаторассеченным листом и ярким пятнистым покрывалом соцветия. Обитают они в затененных местах под пологом тропических лесов и на открытых участках, в саваннах, на низменностях и в горах, поднимаясь до высоты 2500 м в районах Гималаев. Сауроматум капельный (S. guttatum) - декоративное сапромиофильное растение (табл. 64). Во время цветения перед раскрытием пыльников придаток початка соцветия издает резкий, неприятный запах, привлекающий мух-опылителей.

Таблица 64. Аронниковые: 1 - монстера деликатесная (Monstera deliciosa), оранжереи Ботанического института АН СССР в Ленинграде; 2 - аронник удлиненный (Arum elongatum), Крым, г. Аюдаг; 3 - сауроматум капельный (Sauromatum guttatum), Крым, Никитский ботанический сад; 4 - женское соцветие ариземы темно-красной (Arisaema atrorubens), передняя часть покрывала удалена, парк Ботанического института АН СССР в Ленинграде
Именно у этого вида появлению запаха предшествует резкое, буквально в течение дня повышение концентрации свободных аминокислот в 20 раз, что, возможно, и определяет резкость, быстроту появления и кратковременность запаха.
Большинство видов рассматриваемой группы родства - гигрофиты, обитающие во влажных затененных местообитаниях, но есть в подсемействе и растения настоящих пустынь. К ним относятся виды рода эминиум (Eminium), распространенного преимущественно в Ирано-Туранской флористической области. Эминиумы быстро развиваются, цветут и плодоносят весной в короткий период дождей. С прекращением дождей надземная часть растений отмирает, и они проводят большую часть года в состоянии клубня в почве, как обычные эфемероиды геофиты. На фоне светлых красок наших среднеазиатских песчаных пустынь резко выделяется темно-фиолетовыми, почти черными покрывалами соцветий эминиум Лемана (Е. lelimannii). Соцветие его устроено по типу ловушки, и видоизмененные стерильные цветки, занимающие весь промежуток оси початка между женскими и мужскими цветками, вероятно как и у аронников, удерживают насекомых-опылителей в зоне женских цветков. Привлекает же насекомых не только интенсивная окраска внутренней поверхности покрывала, но и отвратительный запах, исходящий от придатка початка. По этим признакам легко узнать способ опыления растения - это уже известная нам сапромиофилия. Зоохория - обычный путь распространения его плодов - сочных белых ягод. Придаточные корни, отходящие от клубней, служат у эминиума хранилищами влаги, иногда ее используют пастухи для утоления жажды. В самих клубнях содержится до 30% крахмала и найдены алкалоиды. Эминиум Альберта (Е. alberti, рис. 274) - горное растение, обитающее в Средней Азии на высоте 500- 1800 м над уровнем моря, декоративное, но ядовитое. Темно-фиолетовое покрывало и причудливо трижды пальчаторассеченные листья придают ему необычность и своеобразие. Произрастает эминиум Альберта обычно среди кустов миндаля в эфемеретумах вместе со злаками. Как декоративные растения эминиумы используются сравнительно мало. Из покрывала их соцветий получают краску, клубнями эминиума Альберта травят мышей и даже крупных хищников. Содержание алкалоидов в клубнях этого растения сравнительно высоко (до 0,5%), и, по-видимому, многие из этих алкалоидов ядовиты.
В подсемействе аронниковых есть ряд небольших родов, распространенных в Неотропическом флористическом царстве. Это южнобразильский род спатикарпа (Spathicarpa), представители которого отличаются своеобразными соцветиями с почти полным срастанием покрывала и початка, что и отражено в названии рода. Интересен род астеростигма (Asterostigma), его вид - астеростигма Лушната (A. luschnathianum) - травянистое растение с характерными для рода звездчатыми рыльцами (рис. 275), распространено в горных районах Бразилии и зовется там "харарака" за сходство пятнистых черешков ее листьев с расцветкой гремучей змеи (харарака - гремучая змея). Небольшие клубни этого растения ценятся как противоядие от змеиных укусов.
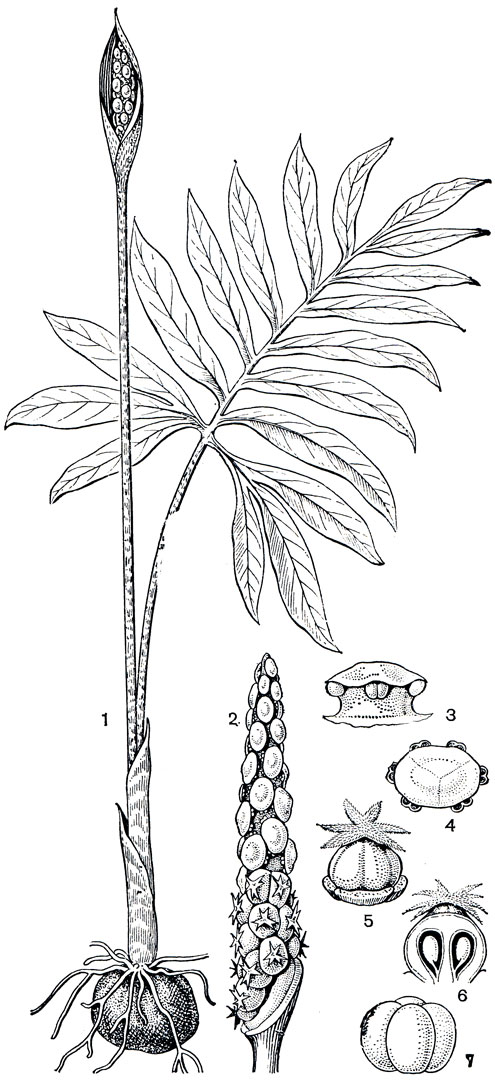
Рис. 275. Астеростигма Лушната (Asterostigma luschnathianum): 1 - общий вид; 2 - початок (покрывало срезано, внизу - женские, вверху - мужские цветки); 3 - мужской цветок (синандрий), вид сбоку; 4 - то же, поперечный разрез; 5 - гинецей (вид сбоку); 6 - то же, продольный разрез; 7 - плод
К подсемейству аронниковых принадлежит ряд небольших, четко обособленных средиземноморских родов. Среди них - олиготипный род аризарум (Arisarum). Его представитель аризарум обыкновенный (A. vulgare) - травянистое растение с яйцевидными или цилиндрическими клубнями, нередко ветвящимися корневищами и стреловидно-яйцевидными листьями. Изогнутые соцветия несут в нижней части 3-6 женских цветков и непосредственно над ними около 40 рассеянно распределенных мужских цветков, каждый из которых состоит из 1 тычинки. Соцветия устроены по типу ловушки и привлекают насекомых отвратительным запахом, сразу после опыления исчезающим. Однако приспособления растения, заставляющие насекомых посетить нижнюю часть соцветия, где расположены женские цветки, существенно отличаются от того, что мы видели у аронников. Здесь уже нет ни скользких сосочков на внутренней поверхности покрывала, по которой опылители соскальзывали к основанию початка, ни стерильных цветков, препятствующих насекомым выйти из соцветия. Специфические приспособления в соцветиях аризарумов рассчитаны на оптический обман насекомого и заключаются в своеобразном распределении темных и светлых участков окраски покрывала, дезориентирующем находящихся в соцветии насекомых. К. Фэгри и Л. ван дер Пэйл (1966) не без основания называют такие соцветия оптическими ловушками.
Насекомые-опылители аризарума обыкновенного, привлеченные запахом, подлетают к растению, проникают в его соцветие и, пытаясь выйти, ползут на свет. Но самыми освещенными участками оказываются места в нижней части соцветия, наиболее удаленные от выхода. Обманутые световым эффектом, насекомые ползут вниз и, безуспешно пытаясь выйти из соцветия через светящиеся прозрачные полосы ("окна") в нижней части покрывала, ползают по початку, опыляют цветки и уносят на поверхности тела новые порции пыльцы. После опыления одногнездная завязь аризарумов, несущая несколько ортотропных семязачатков, развивается в ягодообразный нераскрывающийся плод, тонкий околоплодник которого придает ему сходство с коробочкой. На сравнительно крупных семенах развиваются большие мясистые строфиоли, играющие определенную роль в распространении семян животными. Такого же типа мясистые придатки развиты и у семян амброзинии Басса (Ambrosinia bassii) - единственного вида средиземноморского рода амброзиния, имеющего разорванный ареал (Корсика, Сардиния, Калабрия, Алжир). Это небольшое (высотой 5-10 см) растение с клубнем и 2- 3 яйцевидными листьями, имеет совершенно необычно устроенное соцветие (рис. 276). Помимо того, что расположенные в 2 ряда мужские цветки размещены на отдельном выросте початка, единственный женский цветок изолирован от мужских перегородкой, образованной выростами початка, делящей полость внутри покрывала на две камеры. В одной из них расположен женский цветок, в другой - початок с мужскими цветками. Кроме того, растение страхуется от самоопыления и своеобразным положением соцветия. В процессе развития соцветие отклоняется от растения и, наконец, располагается на поверхности земли так, что сверху оказывается камера с женским цветком, внизу - с мужскими, причем их пыльники повернуты к земле. Проникновение пыльцы к женскому цветку (снизу вверх) исключается, даже если бы и рыльца, и тычинки функционировали одновременно. Такой тип строения и размещения соцветия полностью предотвращает самоопыление. Но в естественных условиях и в культуре амброзиния очень слабо плодоносит, а в отдельных районах вообще не дает плодов, хотя искусственное опыление, даже пыльцой из собственного соцветия, оказывалось эффективным. Все это позволило предположить, что амброзиния "потеряла" своих опылителей. Если это так, то случай с амброзинией еще раз подтверждает односторонность узкой специализации. Полное предотвращение самоопыления - прогрессивное приспособление, но при исчезновении опылителей оно обернулось у амброзинии невозможностью вообще сформировать плоды. В эволюционном плане редукция у амброзинии женского соцветия до одного цветка, размещение мужских цветков на другой оси, изолированность женской и мужской частей соцветия могут рассматриваться как ступень к формированию однополых соцветий и перехода к двудомности.
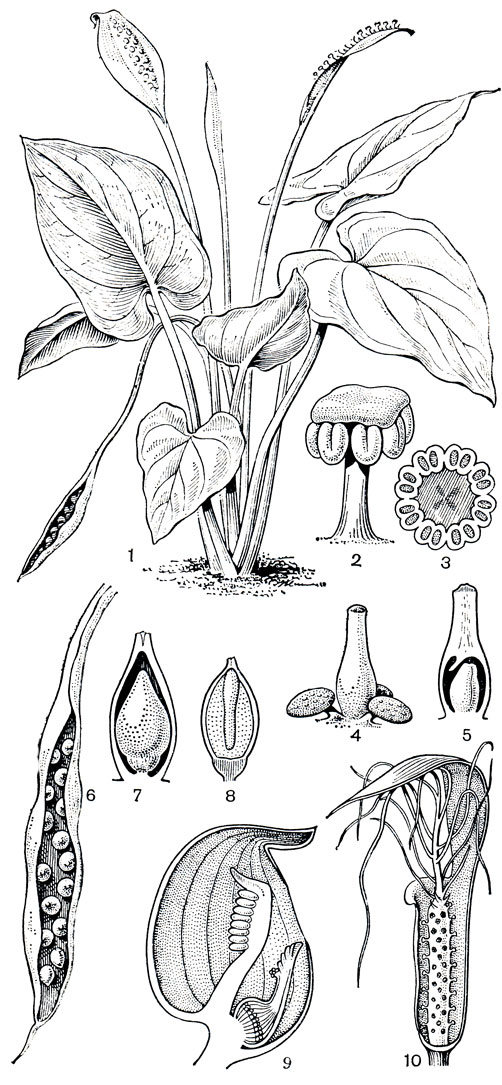
Рис. 276. Собственно аронниковые. Спатикарпа стрелолистная (Spathicarpa sagittifolia): 1 - общий вид; 2 - мужской цветок; 3 - поперечный разрез синандрия; 4 - женский цветок; 5 - продольный разрез гинецея; 6 - покрывало с початком в стадии плодоношения; 7 - плод; 8 - продольный разрез семени. Амброзиния Басса (Ambrosinia bassii): 9 - соцветие, часть покрывала удалена. Аризема Смитинанда (Arisaema smitinandii): 10 - соцветие
На чрезвычайно интересных во флористическом отношении Сейшельских островах географически обособлен от других аронниковых эндемичный монотипный род протарум (Рrоtarum). Его единственный вид протарум сейшельский (P. sechellarum) обитает на острове Маэ среди зарослей пальм. Это небольшое клубненосное растение с рассеченной пластинкой листа и однополыми голыми цветками; впечатление околоцветника в женских цветках создают стаминодии, похожие на части околоцветника. Тычинки соединены в синандрии. Придаток початка, пока еще не отделенный от мужских цветков, образуется слиянием оси соцветия со стерильными цветками.
Примерно половина всех видов подсемейства аронниковые принадлежит роду аризема (более 100 видов), распространенному преимущественно в тропической и субтропической зонах Старого Света, лишь 5-6 видов ариземы - аборигены Нового Света и обитают в атлантическом районе Северной Америки. Ариземы - декоративные травы, развивающие обычно 1 - 4 листа с 3-9-рассеченной пластинкой; стебель подземный, в виде клубня. Соцветия ариземы - оптические ловушки, в очертании похожие на изящный, расширяющийся кверху цветок (один из характернейших признаков рода), причем у многих видов початок или покрывало оканчивается хвостовидно оттянутым образованием, иногда почти нитевидным. Было отмечено, что хвостовидные окончания частей соцветия чаще всего встречаются у сапромиофильных растений. Это натолкнуло на мысль, что структура подобного рода играет определенную роль в процессе опыления. Л. ван дер Пэйл (1953), отметив склонность мух опускаться на свободно подвешенные предметы (вспомним эффект элементарнейшей ловушки для мух - подвешенные полоски липкой бумаги), предположил, что аналогично и хвостовидные окончания частей соцветия могут привлечь мух-опылителей; эти окончания к тому же являются еще и носителями запаха (осмофорами), однако выработка запаха у ариземы, в отличие от аронников, аморфофаллуса, сауроматума и других, не связана с взрывным увеличением интенсивности метаболизма. Запах соцветий аризем менее сильный и ощущается дольше. Чтобы привлечь на этот запах определенный тип насекомого-опылителя, орган запаха, как предполагает немецкий ученый С. Фогель (1954), должен быть выдвинут далеко за пределы соцветия, что и достигается хвостовидным вытягиванием окончаний початка или покрывала. У многих видов аризем женская и мужская части соцветия разобщены полностью и находятся на разных растениях и эта двудомность предотвращает самоопыление. Переход к двудомности осуществился у аронниковых в самой специализированной группе, но и у них двудомность еще не абсолютна и в зависимости от условий питания и развития растения возможен возврат к однодомности. Некоторые же растения имеют несколько форм, отличающихся по полу. Японский ботаник М. Хотта (1971) указывает, что японская аризема Негиши (A. negishii) может существовать в 3 формах: молодые и небольшие растения - мужские; крупные растения - женские и примерно промежуточные растения - обоеполые.
К двудомным растениям принадлежат также виды ариземы, естественно произрастающие в долинных смешанных и лиственных лесах советского Приморья: аризема амурская (A. amurense, рис. 277) и аризема японская (A. japonicum).
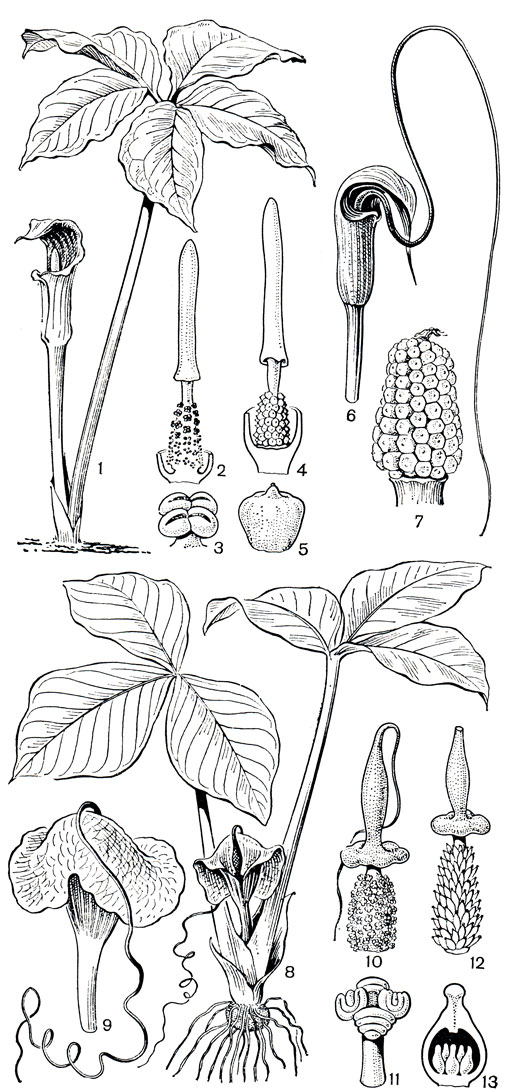
Рис. 277. Ариземы. Аризема амурская (Arisaema amurense): 1 - общий вид; 2 - соцветие мужского растения (покрывало удалено) с мужскими цветками; 3 - тычинка; 4 - соцветие женского растения (покрывало удалено) с женскими цветками; 5 - женский цветок - гинецей. Аризема Тунберга (A. thunbergii): 6 - соцветие с нитевидным придатком; 7 - соплодие. Аризема Гриффита (A. griffithii): 8 - общий вид; 9 - соцветие; 10 - соцветие мужского растения (покрывало удалено) с мужскими цветками в нижней части и фигурным придатком, оканчивающимся нитью; 11 - мужской цветок; 12 - соцветие женского растения (покрывало удалено) с женскими цветками в нижней части; 13 - продольный разрез женского цветка (гинецея)
К двудомным растениям относится и аризема единокровная (A. consanguineum), распространенная в Китае, в провинции Юньнань. У этого вида в процессе жизни существенно меняется форма листа, и трудно представить, что тройчатый лист ювенильного растения и пальчаторассеченный на 15-19 частей лист взрослого экземпляра принадлежат одному и тому же растению. Его соцветие - типичная оптическая ловушка с нитевидно вытянутой верхушкой и вертикальными светлыми полосами в нижней половине покрывала.
Совершенно своеобразную группу гидрофитов образуют 2 близких палеотропических рода: лагенандра (Lagenanclra) - эндемик Индийской флористической области, включающий 5-6 видов, произрастающих преимущественно в стоячих водах болот и по берегам рек, и род криптокорина (Cryptocoryne), распространенный в Индо-Малезийском подцарстве. Его представителей можно встретить в тропической зоне Азиатского континента, на острове Шри-Ланка, на островах Малайского архипелага и в Новой Гвинее. Этот род включает более 50 близких видов, преимущественно земноводных, но развивающихся как плавающие, погруженные и наземные растения. Криптокорины небольшие, цельнолистные, корневищные травы, нередко погруженные растения с яркими соцветиями, возвышающимися над поверхностью воды. Сейчас уже во всем мире виды криптокорины разводят как декоративные аквариумные растения. Преимущественно водный образ жизни способствовал развитию у криптокорины ряда специфических особенностей, одной из них является своеобразное строение их соцветий. Почти все покрывало соцветия преобразовано в длинную узкую трубку с несколько вздутой нижней частью, внутри которой находится короткий початок (1-3 см). Трубка водонепроницаема и ведет водновоздушный образ жизни: ее нижняя часть погружена в воду, верхняя - надводная. Благодаря такому положению початок защищен от намокания, хотя и находится ниже уровня воды. Открытый во время цветения верхний конец труокн позволяет насекомым проникать в соцветие и опылять его цветки. В нижней части початка расположены 6 женских цветков, образующих внешний круг из 6 сросшихся гинецеев, внутренний круг женских цветков преобразован в стерильные ароматические подушечки - осмофоры. Выше по початку после зоны стерильной оси расположены многочисленные мужские цветки (их может быть более 100), каждый из них состоит из 2 тычинок. Оканчивается початок коротким придатком. Нижняя часть трубки покрывала, в которой находится початок, отделена от остальной части клапаном. До цветения этот клапан и верхушка трубки плотно закрыты и открываются лишь непосредственно перед началом женской фазы цветения. Привлеченные цветом покрывала и запахом, исходящим от ароматических подушечек, насекомые проникают в соцветие, но обратно уже выйти не могут (клапан открывается только вовнутрь) и остаются пленниками соцветия. Если во время женской фазы цветения опыление цветков произошло, понижается упругость трубки и клапана, он подвядает и насекомые могут выбраться наружу. В противном случае им можно будет выйти на 2-3 дня позже. Мужская фаза цветения начинается на 3-4-й день. В это время раскрываются пыльники и слизистые скопления пыльцы приклеиваются к ползающим по трубке насекомым, они и переносят эту пыльцу на цветки других соцветий. Просмотр насекомых в гербарных образцах криптокорин позволил монографу рода чешскому исследователю Карелу Ратаи (1975) установить, что их соцветия посещают мелкие насекомые, преимущественно из отрядов двукрылых, перепончатокрылых и жуков (божьих коровок).
Развитие семян у криптокорин относится к тому своеобразному типу вивипарии, который мы уже рассматривали у тифонодорума. Развитие зародыша в семени криптокорины реснитчатой (С. ciliata) исследовал немецкий морфолог Карл Гебель (1931). Почечка зародыша у этого вида, развиваясь внутри семенной кожуры, образует множество (30-40) зеленых листовых зачатков (рис. 278). Семядоля развивает гаусторию, внедряющуюся в эндосперм, и поставляет почечке необходимые для развития вещества. К моменту раскрывания коробочки и освобождения семян от плода в них уже полностью сформирован всход. Попавшее в воду семя очень непродолжительное время плывет по течению, но, как только от него отделяется семядоля, теряет плавучесть, опускается на дно и прорастает. Примерно через год развившееся из этого семени растение уже может образовать соцветие. В естественных условиях, однако, криптокорины исключительно редко размножаются семенами, более обычно у них вегетативное размножение корневыми отпрысками и корневищами. Криптокорины растут обычно в затененных местах по рекам и ручьям, протекающим под пологом дождевых лесов. Они адаптированы к недостатку света и при падении освещенности способны менять окраску листьев от ярко-зеленой, оливковой, коричневатой до интенсивно-красной. На хорошо освещенных отмелях рек, как указывает К. Ратаи, криптокорины развивают множество ярко-зеленых листьев, и преобладают в этих местах виды, предпочитающие наземный образ жизни (криптокорина реснитчатая, криптокорина Вендта - С. wendtii - рис. 279 - и др.). В небольших реках под густой тенью лесного полога обитает криптокорина сердцевидная (С. cordata), обычно развивающаяся как погруженное растение и в затененных глубоких местах образующая только красные немногочисленные листья.
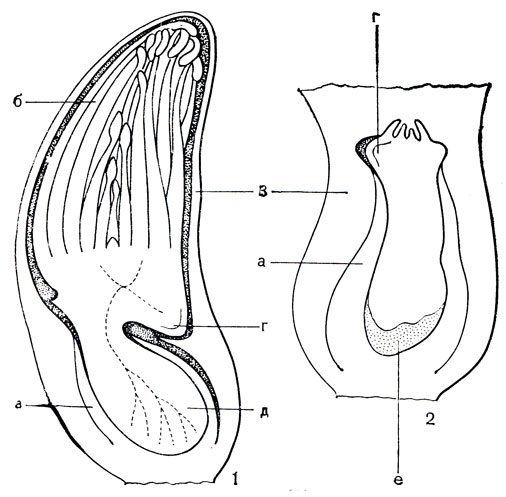
Рис. 278. Развитие зародыша внутри семени у криптокорины реснитчатой (Gryptocoryne ciliata): 1 - зародыш со множеством листовых зачатков; 2 - начальная стадия образования зачатков, а - внутренний интегумент, б - зачаток листа, в - наружный интегумент, г - корешок зародыша, д - семядоля, е - эндосперм
В последнем, наиболее специализированном подсемействе пистиевых (Pistioideae) всего один вид - пистия телорезовидная, или водный латук (P. stratiotes). Это пантропическое плавающее растение с укороченным стеблем, несущим нежные серо-зеленые листья и множество перистых плавающих корней (рис. 279). Листья пистии лопатовидные с наибольшей шириной на конце и несколько суженные к основанию, имеют ряд специальных приспособлений для жизни в плавающем состоянии. Почти параллельные боковые жилки сверху вдавлены, отчего лист кажется гофрированным, но на нижней поверхности они выступают в виде ребер - мощных в основании листа и сходящих на нет к его концу. Такая конструкция придает устойчивость листу, а хорошо развитая воздухоносная ткань увеличивает его плавучесть. Короткие сероватые волоски защищают лист от намокания, выполняя роль водоотталкивающей ткани. Вода, попадая на такой лист, каплями скатывается, не смачивая поверхность листа. Среди скученных на стебле листьев едва заметны зеленоватые волосистые соцветия, обычно не превышающие в длину 2 см. Чуть больше сантиметра початок находится внутри покрывала, неполной перетяжкой отделяющего единственный женский цветок, завязь которого несет многочисленные семязачатки, от 2-8 мужских цветков со сросшимися в синандрии тычинками. Это специализированное соцветие по строению сходно с соцветием амброзинии Басса, но существенно отличается тем, что после раскрывания пыльников пыльца падает непосредственно на рыльце и происходит самоопыление. Несмотря на отсутствие или ограниченность перекрестного опыления пистия обильно плодоносит, развивает полноценные семена и успешно размножается семенным путем. Но значительно быстрее размножается она вегетативно, с помощью столонов, развивающихся в пазухах низовых листьев. На концах столонов возникают новые особи, в свою очередь образующие новые столоны, и так далее. Это небольшое декоративное растение в естественных условиях ведет себя крайне агрессивно и название "злостный сорняк" не полностью отражает степень вреда, причиняемого и природе и человеку этим безудержно размножающимся растением. За короткий промежуток времени пистия может покрыть полностью зеркало воды небольшого водоема и практически обрекает его на исчезновение. Покров из пистии значительно увеличивает расход воды из водоема, так как на ее транспирацию затрачивается несравненно больше воды, чем на испарение с открытой водной поверхности. Она изменяет газообмен в водоеме и подготавливает все условия для полного зарастания его и заболачивания. В отдельных районах десятки километров водной по-верхности покрыты пистией, сотни гектаров заняты этим растением. Скопления пистии затрудняют судоходство, она сильно засоряет посевы риса и таро, нередко сводя на нет затраченный труд. Она создает подходящие условия для жизни москитов и способствует их размножению. Пистия может быстро распространиться на большие расстояния благодаря переносу птицами не только семян, но и вегетативных частей растения. В Африке она успешно расселяется течением рек, передвигаясь на большие расстояния вместе со скоплениями других растительных остатков. Сходным образом вегетативные части пистии переносят реки Индии и Южной Америки. Совсем недавно это настоящее тропическое растение стало осваивать водоемы Дании, быстро развившись в них в необычно жаркое лето 1976 г. Однако датский климат не лучший вариант для семенного размножения пистии. Как установили недавно голландские ученые, семена пистии прорастают после небольшого периода покоя (в несколько недель), но при температуре не ниже 20° С. При хорошей освещенности семена могут прорастать и в воде. Они сохраняют жизнеспособность при падении температуры до 4° С, а также при непродолжительном воздействии отрицательных температур (в пределах - 5° С). Но если даже семенное размножение пистии будет затруднено в водоемах Дании, то ведь это не единственный способ ее размножения. Способствовал расселению пистии и человек; ее разводили как лекарственное растение и в народной медицине использовали для лечения многих болезней. Пистию разводят как аквариумное декоративное растение. Ее можно также использовать на корм и для производства удобрений, однако пока еще не разработана технология, обеспечивающая экономическую выгоду этого производства.
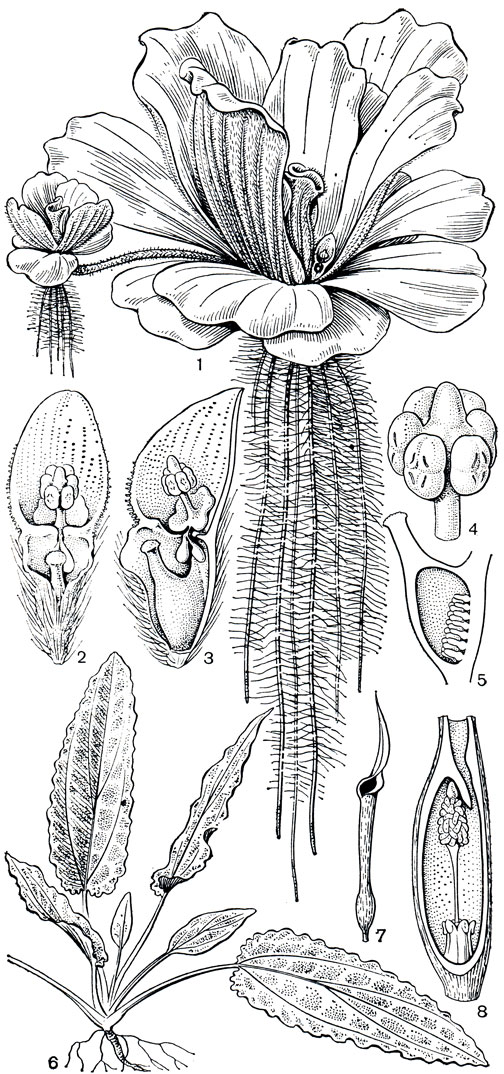
Рис. 279. Водные аронниковые. Пистия телорезов и дна я, или водный латук (Pistia stratiotes): 1 - общий вид; 2 - соцветие (вид спереди); 3 - то же (вид сбоку), часть покрывала удалена; 4 - мужская часть соцветия; 5 - женская часть соцветия - продольный разрез через женский цветок (гинецей). Криптокорина Вендта (Cryptocoryne wendtii): 6 - общий вид; 7 - соцветие; 8 - нижняя часть соцветия (часть покрывала удалена), женские цветки отделены от расположенных вверху мужских цветков узкой стерильной осью соцветия
Семейство рясковые (Lemnaceae) (Т. Г. Леонова)
Это семейство включает 6 родов и около 30 видов, встречающихся на всех континентах. Наиболее широко они распространены в Северной и Южной Америке, в Европе, Южной Азии, Южной и Центральной Африке, на юге Австралии, преимущественно в окультуренных районах. Примерно половина видов обитает в тропиках и субтропиках, остальные - в умеренном поясе.
Представители семейства рясковых являются самыми маленькими в мире цветковыми растениями, величина которых редко превышает 1 см. В результате гидрофильной эволюции они достигли крайней степени редукции всех своих органов, поэтому и по простоте строения занимают первое место среди цветковых. Рясковые относят к числу ботанических курьезов или растений-загадок, до сих пор мало изученных в отношении их биологии и систематики. Не разработана также единая морфологическая терминология для обозначения всех органов рясковых.
Рясковые - водные, свободноплавающие, большей частью многолетние, травянистые растеньица; только ряска тропическая, или равноденственная (Lemna aequinoctialis), из тропической Африки считается однолетней. Вегетативное тело рясковых по виду напоминает плавающий лист или слоевище низших растений. Не удивительно поэтому, что до начала XVIII в. ряску относили к водорослям. Только в 1710 г. итальянский ботаник А. Валлиснери впервые обнаружил у ряски микроскопические цветки. В литературе тело рясковых называется по-разному: филлокладий, вайя, щиток, пластинка, фронд, листец и даже стебель или лист. Наиболее удачным представляется термин "листец", поскольку он лучше других соответствует внешнему облику растения, а также происхождению его тела, которое после работ А. Энглера (1889) большинством ботаников рассматривается как особая структура, не дифференцированная на листья и стебель ("листоветвь"). Листецы рясковых одиночные, или же они соединены в небольшие группы по 2 или более либо в цепочки короткими или удлиненными ножками, образованными суженной частью листеца. Они представляют собой симметричную или асимметричную, большей частью зеленую пластинку, плоскую или уплощенную, реже сильно выпуклую с брюшной стороны. По форме листецы могут быть почковидными, округлыми, эллиптическими, ланцетными и линейными (у ряски - Lemna и многокоренника, или спироделы, - Spirodela) или шаровидными и овальными (у волъфии - Wolffia). Они состоят в основном из паренхимных клеток хлоренхимы, разделенных большими межклеточными полостями, заполненными воздухом или другим газом, что обеспечивает плавучесть растения. Часто присутствуют рафиды и друзы оксалата кальция, а также красные или коричневые (иногда те и другие сразу) пигментные клетки, окрашивающие весь листец (у вольфии и у цветущей ряски) или только его нижнюю часть (многокоренник) в коричневые или красно-фиолетовые цвета. Проводящая система у рясковых практически отсутствует, за исключением многокоренника, у которого в корнях имеются трахеиды. Корни отсутствуют или слабо развиты и не достигают грунта. Они простые, с корневым чехликом, в числе 1 или нескольких отходят от брюшной поверхности листеца. Проксимальная (ее иногда называют базальной) часть листеца ряски и многокоренника расщеплена двумя боковыми кармашками, которые называют также почечными, так как в них закладываются вегетативные почки, дающие начало дочерним листецам при вегетативном размножении. Иногда в одном из кармашков развивается соцветие, вначале окруженное покрывалом. У вольфии, волъфиеллы (Wolffiella), волъфиопсиса (Wolfiopsis) и псевдоволъфии (Pseudowolffia) в этой части листеца имеется только один базальный кармашек, служащий исключительно для вегетативного размножения. Соцветие у них лишено покрывала, или иногда оно заключено первоначально в рудиментарное пленчатое покрывало; расположено соцветие в специальной цветковой ямке (или в двух ямках) на дорсальной поверхности листеца.
Соцветие рясковых сильно упрощено до 1-2, редко 3 (у многокоренника) мужских цветков и 1 женского. Большинство современных ученых склоняются к мнению, что у рясковых мы имеем дело с соцветием, представляющим собой сильно редуцированный початок ароидных, которые считаются предками рясковых. Эмбриологическими исследованиями индийского ботаника С. Махешвари (1954, 1956, 1958) установлено, что многокоренник обыкновенный (Spirodela polyrhiza) можно считать промежуточным звеном между ароидными и другими родами рясковых. Околоцветник у рясковых отсутствует. Мужские цветки состоят из 1, редко 2 тычинок. Пыльники 2-гнездные, раскрывающиеся поперечно, или одногнездные, вскрывающиеся вдоль верхушечной пигментной линии; прикреплены они к тонкой или веретеновидной тычиночной нити своим основанием. Оболочка пыльцевых зерен однопоровая, шиповатая. Женские цветки помещаются между мужскими. Гинецей у них псевдомономерыый, с 1-4, реже 7 базальными семязачатками. Столбик короткий, с рыльцем в виде открытой чаши. Плоды рясковых мешочкообразные, широкоовальные и слабокрылатые или шаровидные, иногда слегка продольно сплюснутые; они нераскрывающиеся или раскрывающиеся продольно, содержат от 1 до 6 семян. Семена крупные, с прямым зародышем, снабжены скудным эндоспермом или лишены его. Они овальные или почти шаровидные, могут быть продольно или сетчато-ребристыми, сетчатыми или гладкими, с губчатым наружным слоем и с уплощенной крышечкой на микропилярном конце.
По своей природе рясковые являются неотеническими формами (А. Л. Тахтаджян, 1943, 1966), произошедшими от предков современного водоплавающего тропического рода пистия (Pistia) из семейства ароидных. Эволюцию тела рясковых внутри самого семейства подробно описывает И. Е. Иванова (1973). Она считает, что вначале проросток предполагаемого предка современных рясковых был равносемядольным, но с нерасходящимися семядолями и замкнутой между ними точкой роста побега, а также со стержневым корнем. Затем он постепенно становился неравносемядольным и приобрел свойственную рясковым асимметрию. Стержневой корень при этом утратился, а вместо него, пробуравив нижнюю семядолю, возник пучок придаточных корней, смещенных к центру листеца и обеспечивающих ему устойчивое положение. Количество корней в дальнейшем уменьшилось до одного, или произошла их полная редукция. Появление двух боковых кармашков объясняется наложением друг на друга листовых пластинок, заключающих горизонтальный конус нарастания, который производит как новые листецы, так и репродуктивные органы. При этом дно кармашков образовано меристемой этого конуса, а их стенки - несросшимися здесь листовыми пластинками, которые в остальной части листеца срослись в его монолитное тело.
Семейство рясковые по строению листецов, положению соцветий и наличию или отсутствию покрывала при них разделяется на 2 подсемейства: наиболее примитивное собственно рясковые (Lemnoideae) и более подвинутое волъфиевые (Wolffioideae).
Подсемейство рясковые характеризуется наличием одного корня или пучка из нескольких корней, двумя боковыми кармашками на листеце, одиночным соцветием, заключенным в пленчатое покрывало и состоящим из двух мужских и одного женского цветка. Представители подсемейства вольфиевых не имеют корней, снабжены одним базальным кархмашком и одной, реже двумя цветковыми ямками на дорсальной поверхности, расположенными вправо и влево от средней линии листеца; соцветие у них одно, реже их 2, каждое состоит из одного женского и одного мужского цветка и лишено пленчатого покрывала. В подсемействе рясковых 2 рода: ряска (Lemna, 9 или более почти космополитных видов) и многокоренник (Spirodela, 4 вида, широко распространенных в умеренном и тропическом поясах обоих полушарий). Собственно рясковые - наиболее крупные растения, хорошо заметные простым глазом. Они зеленые и напоминают маленький плавающий лист. Многокоренник всегда обитает на поверхности воды. Его плоские яйцевидные или обратнояйцевидные, иногда почковидные листецы несут чешуйки на обеих сторонах и снизу часто бывают красными или красновато-фиолетовыми благодаря присутствию пигментных клеток в эпидерме. Встречаются они поодиночке, группами из 2-5 или спирально закрученными цепочками из 8 листецов, соединенных отходящими от их нижней стороны короткими ножками, как это мы видим у многокоренника обыкновенного, известного на всех континентах и на многих крупных островах (рис. 280). Листецы многокоренника длиной 3-10 мм и шириной 1,2-8 мм, на нижней поверхности всегда несут пучок из 2-20 корней, длина которых зависит от глубины водоема и колеблется в пределах 1 -10 см. У многокоренника точечного (S. punctata) иногда развивается только 1 корень. Корни у рясковых выполняют главным образом функцию якоря, предотвращая переворачивание растений. Переплетаясь между собой, корни образуют спутанную массу, облегчающую расселение и увеличивающую устойчивость колонии, что особенно важно при обитании в текучей воде. Питательные вещества из окружающей среды усваиваются всей (у погруженных видов) или только брюшной поверхностью листецов.
Большинство видов ряски также плавает на поверхности воды, но некоторые виды, как, например, ряска тройчатая (L. trisulca), встречающаяся в Северной Америке, Евразии, Африке и Австралии, являются погруженными растениями, т. е. она обитает под водой близ самой ее поверхности, всплывая лишь в период цветения. Листецы ряски также бывают одиночными, или они соединены в группы по 2-10 с помощью отходящих от края более или менее длинных ножек. Ряска тройчатая образует длинные, иногда ветвящиеся, спиралевидные цепочки из 3-22, а иногда и до 50 листецов. Попадая в глубь водоема, где мало света, такая цепочка, вращаясь, снова всплывает. Форма листецов рясок может быть округлой, эллиптической, продолговатой, обратнояйцевндной или ланцетной. Размеры их очень малы. Самые маленькие ряска крошечная (L. perpusilla) из восточной части Северной Америки и ряска мелковатая (L. minuscula) из Северной и Южной Америки имеют в длину 1-2,5 мм и в ширину соответственно 0,7-2 и 0,7-1,5 мм. Длина листеца самого крупного вида - ряски тройчатой - вместе с длинной ножкой составляет 5-20 мм, а ширина - 2,5-5 мм. Большей частью листецы рясок симметричные и плоские. Только ряска горбатая (L. gibba), растущая в Европе, Западной и Южной Азии, в Африке, Северной и Южной Америке, и ряска двусемянная (L. disperma) из Южной Австралии и Новой Зеландии с нижней стороны имеют хорошо заметную выпуклость, заполненную воздухоносными полостями, отчетливо видными на поперечном срезе (рис. 280).
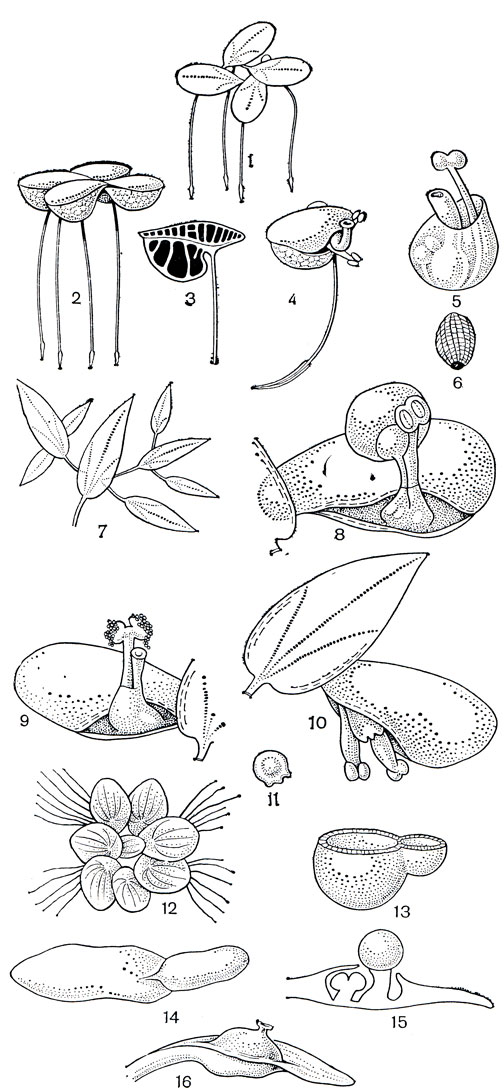
Рис. 280. Рясковые. Ряска малая (Lemna minor): 1 - общий вид цепочки листецов. Ряска горбатая (L. gibba): 2 - общий вид цепочки листецов; 3 - продольный разрез листеца; 4 - цветущий листец; 5 - соцветие, окруженное покрывалом; 6 - плод. Ряска тройчатая (L. trisulca): 7 - общий вид цепочки листецов; 8 - цветущий листец с каплей на рыльце до созревания пыльцы мужского цветка; 9 - цветущий листец в период созревания пыльцы (происходит автогамия); 10 - окончание цветения, появление плода; 11 - плод. Многокоренник обыкновенный (Spirodela polyrhiza): 12 - спирально закрученная цепочка листецов. Вольфия бескорневая (Wolffia arrhyza): 13 - листецы. Вольфиелла языковидная (Wolffiella lingulata): 14 - дочерний листец, появляющийся из кармашка материнского; 15 - соцветие в цветковой ямке; 16 - плод на отмершем родительском листеце
В эпидерме у рясок, как и у многокоренника, имеются красные и коричневые пигментные клетки, паренхима содержит рафиды. У рясок развивается только 1 корень, который у ряски крошечной не превышает 3 см, а у ряски горбатой может достигать в длину 16 см. Ряска тройчатая иногда лишена корней. Соцветие в подсемействе рясковых вначале окружено рудиментарным пленчатым покрывалом, которое образует подобие мешочка (ряска тройчатая, многокоренник обыкновенный), или чаще покрывало представляет собой открытое ложе с загнутыми вверх краями. В соцветии, как правило, развиваются 2 мужских цветка, но может быть 1 или 3. Величина тычинки немногим более 1 мм. Двугнездные пыльники прикреплены к уплощенным нитям и снабжены коническим заостренным связником. Вскрываются они почти поперечной щелью, расположенной на дне специальной бороздки; к этому времени перегородка между пыльниками исчезает и они становятся одногнездными. Женский цветок редко превышает в длину 1 мм. Завязь яйцевидная, одногнездная, с 1-6 семязачатками; столбик короткий и тонкий, рыльце расширено в форме неглубокой воронки или чаши с неровными краями. Цветение в семействе рясковых не укладывается ни в какие известные для цветковых растений ритмы, и наблюдалось оно настолько редко, что специально регистрировалось. Так, в Финляндии, например, с 1895 по 1947 г. отмечено 33 случая цветения рясок; в Польше с 1679 по 1959 г. цветение ряски зафиксировано только 2 раза. В Америке цветущий многокоренник за последние 200 лет наблюдали не более 20 раз. По сообщению A. Л. Тахтаджяна, в 1932 г. в Армении в озерах Дорийской нагорной равнины наблюдалось массовое цветение ряски. Для средней полосы России за период с 1814 по 1967 г. в печати появились сообщения лишь о 25 находках цветущих рясок. Однако о массовом цветении ряски тройчатой, ряски горбатой и единичном ряски малой (L. minor) , в прудах города Новгорода в 1968 г. сообщает И. Е. Иванова (1970), а Г. А. Лукиной (1977) в бассейне реки Волги удалось наблюдать ежегодное цветение ряски с 1972 по 1975 г. Возможно, что цветение рясковых не всегда замечается ботаниками из-за микроскопических размеров цветков.
Цветут ряски в июне-августе, иногда в сентябре. К началу цветения более или менее длинные цепочки разрываются на более короткие, состоящие из 4-5 листецов у ряски горбатой и из 2-4 листецов у ряски тройчатой, у которой эти короткие цепочки всплывают на поверхность. Перед началом цветения в цепочке осуществляется также постепенное морфологическое преобразование листецов из вегетативных в специальные цветущие и происходит это следующим образом (рис. 280). У ряски тройчатой, например, исходный вегетативный листец утончается, бледнеет, его "верхушка" загибал ется вниз. Вначале он производит подобные себе заостренные листецы, которые отличаются лишь меньшими размерами и короткими ножками. Затем образуются еще более мелкие темные и плотные листецы с едва различимыми ножками. И наконец, в кармашках последних развиваются цветущие темно-зеленые листецы с закругленной "верхушкой", незаметными ножками и глубоко просвечивающим цветковым кармашком с одной стороны. У ряски горбатой цветущие листецы становятся оливково-пурпурными, так что островки их хорошо заметны на изумрудном фоне нецветущих особей.
К началу цветения листецы всегда располагаются на поверхности воды и имеют более развитые, чем у вегетативных, воздушные камеры. Каждый листец за свою жизнь производит только одно соцветие, которое у рясок развивается всегда лишь в одном из боковых кармашков, чаще всего в левых (перевернутая симметрия). Реже соцветие образуется в правых (зеркальная симметрия) кармашках цветущих листецов. У многокоренника соцветия могут закладываться в обоих кармашках сразу. С момента заложения цветущего листеца до окончания процесса цветения проходит около 20 дней.
Для всех рясковых характерна протогиния, т. е. вначале созревает рыльце женского цветка, затем тычинка одного из мужских цветков и после этого тычинка второго цветка. Процесс цветения у рясковых проследим на примере ряски тройчатой. Из кармашка цветущего листеца сначала появляется столбик, приподнимающийся над водой и изгибающийся так, что устье воронки рыльца обращается в сторону самого листеца. В устье рыльца появляется крупная водянистая капля, далеко выдвигающаяся над поверхностью листеца и служащая, по-видимому, для привлечения мелких насекомых. При прикосновении капля втягивается внутрь рыльца вместе с попавшей на нее пыльцой. Если опыления не произошло, то через несколько часов капля появляется вновь. Рыльце сохраняет жизнеспособность в течение суток, иногда меньше, или же немногим более двух суток. В этот период происходит перекрестное опыление. Затем рыльце отмирает, превращаясь в массу слизи. У одних растений рыльце остается жизнеспособным лишь до момента созревания пыльцы первого (расположенного ближе к "верхушке" листеца) мужского цветка. У других оно сохраняется до созревания второго, базального, мужского цветка, но тогда положение рыльца меняется - оно поворачивается устьем кверху, образуя под вскрывшимися пыльниками первого мужского цветка подобие плотно прижатой к тычиночной нити чаши, в которую легко попадает собственная пыльца и может произойти самоопыление (рис. 280). Тычинка второго мужского цветка выдвигается из кармашка цветущего листеца спустя 24 ч (у ряски горбатой через 11-12 ч)
после появления первого. К моменту рассеивания ее пыльцы пыльники тычинки первого цветка закрываются, а сама тычинка опускается на воду и становится прозрачной. Рыльце в это время уже разрушено и пыльца второго цветка может опылять лишь соседние цветущие экземпляры в других цепочках ряски. Тычинка первого цветка функционирует около 60 ч, второго - около 90 ч. Вначале в зрелых пыльниках обнаруживается "энтомофильная" слабошиповатая ярко-желтая пыльца, которая выступает из пыльников в виде плотно сцепленных влажных комочков. Эти комочки отрываются и падают на поверхность сплавин, по которым часто снуют мелкие насекомые (водяные клещики, тли и др.). В пользу насекомоопыляемости у рясок говорит также окрашивание цветущих листецов в темно-зеленые, оливковые и пурпурные цвета, объединение их в сплавины, характерный поворот рыльца. Через 12-24 ч после вскрывания пыльников пыльца приобретает "анемофильный" облик - тускнеет, становится блеклой и легко сыпучей. В это время она переносится воздушными течениями на большие расстояния. Таким образом, вначале рясковые являются насекомоопыляемыми, а затем ветроопыляемыми растениями. Если перекрестного опыления не произошло, то у них имеет место самоопыление. После увядания рыльца и столбика становится хорошо заметным молодой плодик, находящийся внутри кармашка в виде красноватого вздутия. Плоды рясок чуть больше макового зернышка и хорошо видны невооруженным глазом (рис. 280). Они овальной формы, снабжены зелеными крыловидными выростами по бокам и килем на нижней поверхности. Киль наряду с большими межклетниками и тканями облегчает плавание плода, которое продолжается на поверхности воды всего одни-двое суток. После этого при температуре воды не ниже 18° С семена прорастают в новое растение. Плодоношение, как и цветение, наблюдается в течение июня-августа. Часть семян, особенно образовавшихся в конце лета, не прорастает в холодной воде водоемов и в сентябре-октябре погружается на дно, где перезимовывает, прорастая в мае следующего года.
Плоды семейства рясковых переносятся на большие расстояния на лапках водоплавающих и болотных птиц. Они могут разноситься также и водой. Преобладает у рясковых, однако, расселение вегетативными особями, так как плоды образуются редко. Взрослые растения (и части сплавин) распространяются медленно текучими водами, лягушками и тритонами, к телу которых они прилипают, но преимущественно птицами. Многие дикие утки поедают ряску и многокоренник. В это время растения прилипают к их головам, шеям, вместе с грязью - и к лапкам. Они не погибают в течение 12, а иногда и до 22 ч, находясь на открытом воздухе, как, например, ряска малая в опыте Г. Ридли (1930). За это время утки могут улететь на расстояние более 300 км. В Китае, на полуострове Малакка и острове Суматра на рисовые поля, где ряска является сорняком, ее заносят домашние утки. Разносятся рясковые и питающейся ими ондатрой, реже - на ногах лошадей и крупного рогатого скота. По мнению Ридли, европейские ряска малая и ряска тройчатая на многие острова (такие, как Галапагосские, Куба, Ямайка, Пуэрто-Рико, Бермудские, Канарские и Фиджи) были занесены человеком.
Размножаются рясковые главным образом вегетативно с помощью почек, находящихся в почечных кармашках. Эти почки производят новые дочерние листецы по типу розеток все время в одном направлении; такой порядок можно сравнить с завитком. У многокоренника, у некоторых рясок (как и у подсемейства вольфиевых) образование дочерних растеньиц происходит поочередно то из одного, то из другого кармашка материнского листеца; у ряски тройчатой это происходит из двух кармашков сразу, поэтому спираль менее заметна. Вегетативное размножение продолжается с июня по август и осуществляется очень быстро. При этом рясковые удваивают массу своего тела за 1-6 суток, подобно водорослям и грибам, а удвоение количества листецов происходит за 2-3 суток. В течение своей жизни каждое растение производит значительное количество дочерних, которые некоторое время соединяются с материнским с помощью короткой гиалиновой или длинной зеленой ножки в группы или цепочки, затем отрываются и становятся самостоятельными особями.
Перезимовывают рясковые, а также переносят неблагоприятные для роста условия в виде семян (ряска горбатая, ряска крошечная) или обычных листецов, которые к этому времени утолщаются, становятся более округлыми, заполняются тяжелым крахмалом (ряска тройчатая) и оседают на дно. Многие виды, кроме того, производят особые покоящиеся почки, или турионы, которые более устойчивы к крайним условиям. Наиболее специализированы и известны турионы многокоренника обыкновенного. Они представляют собой округлые диски диаметром 2-3 мм, темно-зеленого или пурпурного цвета. Это видоизмененные листецы, но они меньше и толще обычных. Воздухоносные полости у них редуцированы или полностью отсутствуют, а клетки густо заполнены крахмалом, благодаря чему тяжелые турионы погружаются на дно и остаются там до весны без каких-либо изменений. У ряски горбатой и ряски
малой турионы слабо отличаются от вегетативных. Они тонкие, более темные, лишены воздухоносных полостей, заполняются крахмалом и из поверхностно плавающих становятся полупогруженными, что позволяет им избежать вмерзания в лед. Турионы, или покоящиеся почки, иногда называют зимующими почками, что не совсем удачно, так как они могут образоваться не только в конце, но и в начале лета, а также в жарких странах, где не бывает зимы. Они появляются как при низких (ниже 10 °С), так и при высоких (+25 °С) температурах воды. Весной или при благоприятных температурных условиях и достаточном освещении турионы всплывают на поверхность и из них вырастает новое растение.
Подсемейство вольфиевые включает 4 рода: волъфия (Wolffia, 7 видов, широко распространенных в умеренном и тропическом поясах обоих полушарий), волъфиелла (Wolffiella, 5- 8 видов в тепло-умеренных, субтропических и тропических областях Америки, 1 вид в Южной Африке), псевдоволъфия (Pseudowolffia, 3 вида в Северной и Центральной Африке) и волъфиопсис (Wolffiopsis, 1 вид в тропиках Америки и Африки).
Все вольфиевые лишены корней. Часть из них (вольфиелла, псевдовольфия и вольфиопсис) имеет плоский и тонкий листец длиной 2-9 мм и шириной 0,4-5 мм, линейной, продолговатой, широкоэллиптической или дисковидной формы (рис. 280). Эти растения плавают на поверхности воды или близ нее. Виды вольфии, в отличие от остальных, обладают объемным и толстым, иногда уплощенным на дорсальной стороне (рис. 280) листецом шаровидной, овальной или яйцевидной формы; они всегда плавают на поверхности воды. Вольфия - самое маленькое цветковое растение в мире, редко превышающее 1 мм в длину и ширину. В наших широтах обитает волъфия бескорневая (Wolffia arrhiza). Простым глазом это крошечное растение трудно рассмотреть; но колонии его хорошо заметны и напоминают пятна кофе на воде благодаря коричневому пигменту, содержащемуся в клетках эпидермы. Присутствие этого пигмента характерно для всех вольфиевых, друзы же у них не образуются.
Размеры и форма листецов вольфиевых в значительной степени зависят от времени года. В Калифорнии, например, по наблюдению Г. Мейсона (1938), листецы волъфиеллы языковидной (Wolffiella lingulata) имеют максимальные размеры и наибольшее разнообразие форм в зимние месяцы. Ножки у них в это время довольно длинные, и часто дочерние листецы, Прикрепленные к родительскому, образуют крупные колонии, в составе которых Мейсон в январе насчитал до 20 узких и длинных листецов. Летом колонии образуются редко и состоят не более чем из 5 широких и коротких листецов, соединенных короткими ножками. Проводящая система вольфиевых еще более редуцирована по сравнению с рясковыми и сведена в листеце до стеблевого узла и двух элементов типа междоузлия. Зачатки проводящей системы иногда наблюдаются в тычинках.
В отличие от собственно рясковых на дорсальной поверхности листеца вольфиевых имеется всего 1 базальный кармашек в виде щелевидного или конусовидного углубления (рис. 280). Этот кармашек служит для вегетативного размножения, так как в нем закладываются почки, дающие начало только новым листецам. Вход в кармашек обращен к дистальному, обычно приближенному к воде или погруженному концу листеца, что облегчает молодым растениям попадание в воду. У вольфиевых, так же, как и у рясковых, преобладает вегетативное размножение. Цветущие растения встречаются довольно редко, и долгое время ученые считали, что вольфиевые полностью потеряли способность цвести. Лишь в 1935 г. М. Джиарделли обнаружил в Аргентине цветки волъфиеллы продолговатой (Wolffiella oblonga), а в 1937 г. Г. Мейсон нашел в Калифорнии цветущие экземпляры вольфиеллы языковидной, которая цвела с июня до конца августа. Подобно остальным рясковым, цветущие листецы вольфиевых всплывают на поверхность воды. Они также меньше по размерам, чем вегетативные, и в некоторых случаях (как, например, у вольфиеллы) расширяются на дистальном конце, что приспосабливает листец к образованию дорсальной цветковой ямки. На каждом цветущем листеце находится 1 соцветие, только у вольфиопсиса 2 соцветия в двух цветко-вых ямках. Соцветие вольфиевых лишено покрывала и состоит из 1 женского и 1 мужского цветка. Мужские цветки имеют в длину около 0,8 мм. Пыльники у них одногнездные, раскрываются вдоль верхушечной пигментной линии. Женские цветки не превышают в длину 0,5 мм. В их завязи содержится всего 1 семязачаток; столбик короткий и толстый, переходящий в воронковидное рыльце. В соцветии первым развивается женский цветок. Когда созревает рыльце, на нем появляется такая же капля жидкости, как и у ряски и многокоренника; она отсутствует только у видов вольфии. В остальном процесс цветения у вольфиевых происходит так же, как у рясковых. Плод созревает, оставаясь на листеце. Его размеры составляют около 0,5 мм в длину и 0,3 мм в ширину. Вскоре после созревания семя прорастает через щель в крышечке в молодое растеньице, которое высовывается вначале своим дистальным концом и через некоторое время отрывается, становясь свободно плавающим растением. Материнский листец после созревания и прорастания плода погибает.
Разносятся вольфиевые, как и рясковые, в виде листецов, реже плодов водоплавающими птицами, преимущественно утками, а также лягушками и жабами. Иногда крошечные шарообразные или зернообразные листецы вольфии переносятся ветром. Неблагоприятные условия, в том числе и зимние холода, вольфиевые переносят с помощью погружающихся на дно турионов, которые сходны, как, например, у вольфии, по форме и размерам с обычными листецами, но более тяжелые из-за заполняющего их крахмала.
Семейство рясковые представляет собой не только морфологически, но и экологически изолированную группу. Они являются гидрофитами, свободно плавающими на поверхности или же у самой поверхности воды, так что очень маленькая часть листеца выставляется над водой, или же они полностью погружены, всплывая на поверхность лишь во время цветения. Их можно встретить повсюду в лужах, мелких прудах, канавах, запрудах и других хорошо прогреваемых водоемах с пресной стоячей или медленно текучей, богатой органическими веществами водой. Часто рясковые образуют большие скопления (сплавины), сплошь покрывающие поверхность стоячих неглубоких водоемов. Ряска, многокоренник и вольфиелла встречаются вместе с рогозом (Typha), камышом (Scirpus), роголистником (Ceratophyllum), урутью (Myriophyllum) и другими водными растениями. Рост рясковых в течение некоторого времени может продолжаться и вне воды на влажной грязи.
Рясковые служат кормом для диких и домашних уток, а также других водоплавающих и болотных птиц, для рыб, особенно карпа, и ондатры. В сельском хозяйстве их используют в свежем и сушеном виде как ценный белковый корм для свиней и домашней птицы. С недавнего времени рясковые разводят в культуре из-за высокого содержания протеина (до 45% от сухой массы; 45% составляют углеводы, 5% - жиры и остальное - клетчатка), неприхотливости к условиям выращивания и из-за большой продуктивности биомассы. Так, в культуре вольфия бескорневая на площади среды в 1 га образует до 320 кг воздушно-сухой массы. Применяются рясковые и для очистки воды, так как извлекают из нее и запасают в своих листецах азот, фосфор и калий, а также поглощают углекислый газ и обогащают воду кислородом. Особенно перспективна их культура в районах с круглогодичным вегетативным периодом. Употребляются рясковые, по сообщению Э. Ландольта (1980), в пищу и человеком. В Мексике, например, ряску горбатую продают на пищевых рынках. В Таиланде, Бирме и Лаосе волъфию шаровидную (Wolffia globosa) местные жители используют как овощ или приправу под названием "khai-nam" ("водные яйца"). Рясковые были важным пищевым продуктом в Гватемале для древних майя, называвших их "Xima ha" ("водный маис"). В Португалии, США, Африке и Азии некоторые виды ряски являются сорняками рисовых полей, а вольфия бескорневая в Южной Африке и на Мадагаскаре относится к числу опасных сорняков. Для борьбы с ними применяют как химические, так и механические (например, удаление граблями) способы борьбы. Рясковые можно использовать как органические удобрения.
За последние 50 лет рясковые рассматривают как чрезвычайно ценный экспериментальный объект для морфогенетических, физиологических и биохимических исследований благодаря неприхотливости к среде, малым размерам, быстрому росту, относительной простоте строения и преобладанию вегетативного размножения, что позволяет использовать всего один генетически однородный клон на протяжении всего эксперимента. Кроме того, контрольные условия температуры, света и питания рясковым легче обеспечить, чем другим цветковым растениям.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'