Порядок тиссовые (Taxales)
Семейство головчатотиссовые (Cephalotaxaceae) (О. А. Муравьева, В. С. Борхвардт)
Семейство представлено одним родом - головчатотиссом (Cephalotaxus), состоящим всего из 6 видов. Филогенетически головчатотиссовые тесно связаны с только что представленным читателю семейством подокарповых, особенно с примитивными представителями рода подокарп. По некоторым, присущим ему особенностям, это семейство является как бы промежуточным звеном между примитивными подокарповыми и тиссовыми.
В прежние геологические времена головчатотисс был довольно широко распространен в северном полушарии, в настоящее же время встречается только в Азии, а именно: от Северо-Восточной и Восточной Индии до Центрального и Юго-Восточного Китая, острова Тайвань, в Южной Корее и Японии (карта 34). Растения этого рода предпочитают умеренно теплый климат и влажную, тенистую среду, а потому встречаются главным образом в смешанных горных лесах на высоте 300-3300 м над уровнем моря.
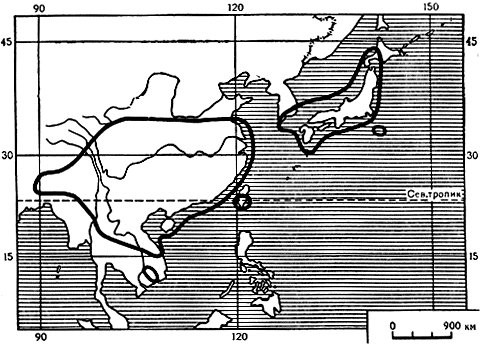
Карта 34. Ареал рода головчатотисс
Головчатотиссовые - вечнозеленые, двудомные, редко однодомные деревья сравнительно небольшой высоты (до 10-15 м) или кустарники. Ветви у них супротивные или мутовчатые. Листья на боковых побегах двурядно расположенные, супротивные или почти супротивные, на вертикальных - расположенные спирально, кожистые, узколинейные, низбегающие по побегу, дорсивентральные, сверху зеленые с ясно выраженной средней жилкой, снизу - с двумя широкими белыми устьичными полосками, более широкими, чем зеленые края листа.
Особенностью семейства является наличие шаровидных (головчатых) собраний микростробилов (отсюда и произошли названия рода и семейства). Эти сложные собрания микростробилов расположены на коротких ножках на побегах последнего года (рис. 223). В шаровидном собрании развивается 6-11 микростробилов, каждый из которых имеет свою кроющую чешую и несет 7-12 микроспорофиллов. К верхушке микроспорофилла прикреплены 3, реже 4 микроспорангия. Микроспоры здесь без воздушных мешков, с тонкой экзиной и толстой интиной.
Мегастробилы собраны в мелкие шишки, развивающиеся по 1-3 в пазухах чешуевидных листьев на верхушках побегов. Шишка состоит из 4-6 пар компактно крестообразно расположенных кроющих чешуй, утолщенных у основания. Каждый элементарный мегастробил, сидящий в пазухе кроющей чешуи, состоит из двух прямых семязачатков. Семязачатки при основании окружены эпиматием в виде воротничка, который по мере созревания семени разрастается и превращается в наружный мясистый покров семени со смоляными полостями. Эпиматий срастается с интегументом, последний становится каменистым, зрелое семя при этом приобретает сходство с плодом-костянкой покрытосеменных. В шишке обычно развиваетется только по одному, редко по 2-3, костянковидному семени (рис. 223). Зрелое семя обычно эллипсоидальное, крупное, 2-3 см длины, красноватое или оливково-коричневое. Зародыш с двумя довольно широкими семядолями, которые при прорастании семени сбрасывают семенную кожуру и фотосинтезируют (надземное прорастание).
Древесина головчатотиссовых не имеет отчетливого разделения на ядро и заболонь, она желтовато- или коричневато-белая. У деревьев, растущих в субтропических областях, годичные кольца плохо заметны, но при выращивании к северу от границы естественного ареала они часто более или менее отчетливые. Смоляные каналы в древесине отсутствуют, иногда имеются лишь в сердцевине. Древесинная парен-хима большей частью диффузная. Трахеиды имеют от 1 до 3 спиральных утолщений. На радиальном срезе в трахеидах ранней древесины окаймленные поры преимущественно однорядные, округлые, вход в окаймленную пору щелевидный, иногда крестообразный, в трахеидах поздней древесины оба отверстия щелевидные. Торус хорошо заметен. Радиальные лучи без лучевых трахеид, большей частью однорядные.
Мезофилл листьев головчатотиссовых дифференцирован на палисадную (дву- или однослойную) и губчатую ткань, клетки последней горизонтально вытянуты от пучка к краям листа. Под пучком имеется небольшой смоляной канал.
Изучением эмбриологии видов головчатотисса занимались многие ученые. Одним из первых был В. И. Арнольди, ботаник из школы известного русского эмбриолога XIX в. И. Н. Горожанкина. На примере изучения головчатотисса Харрингтона (С. harringtonia) ему удалось выяснить, что в марте микроспоры, содержащиеся в микроспорангиях, еще одноядерные, в апреле или мае начинается их рассеивание, продолжающееся около 3 недель. К началу рассеивания микроспор они становятся двуклеточными, так как ядро микроспоры образует более маленькую генеративную клетку (проталлиальные клетки полностью отсутствуют). В таком двуклеточном состоянии пыльцевые зерна попадают на верхушку нуцеллуса. К этому времени семязачатки еще неотчетливо заметны, они имеют вид бугорка, состоящего из недифференцированного нуцеллуса, окруженного интегументом. Только в мае следующего года пыльцевые зерна прорастают на нуцеллусе. Перед проникновением пыльцевой трубки в нуцеллус генеративная клетка делится на сперматогенную клетку и стенную, или клетку-ножку.
Обстоятельная работа, посвященная формированию женского гаметофита у головчатотисса Форчуна (С. fortunei) принадлежит Е. М. Соколовой (1890 г.). По данным этого автора, в начале апреля приблизительно через год после начала развития семязачатка в сформи-ровавшемся женском гаметофите начинается развитие архегониев, и только в конце мая, т. е. спустя примерно 15 месяцев после образования мужского гаметофита, происходит оплодотворение.
Процесс оплодотворения и ранние стадии развития проэмбрио (предзародыша) впервые были изучены в 1900 г. В. И. Арнольди. На женском гаметофите близ микропиле образуется по 2-5, чаще 4 архегония. Они имеют сильно вытянутую форму с оттянутым нижним (халазальным) концом. Шейка состоит из 2-5 клеток, имеется и ядро брюшной канальцевой клетки, которое дегенерирует перед оплодотворением.
Перед самым оплодотворением сперматогенная клетка делится, образуя два обычно одинаковых спермия. При оплодотворении в архегонии вступают не только спермии, но и вегетативное ядро пыльцевой трубки, а также клетка-ножка. Если спермии различны по величине, то с ядром яйцеклетки сливается более крупный.
Вскоре после оплодотворения в зиготе образуются 4 свободных ядра. В стадии 16 ядер начинается обособление клеток, которые затем располагаются в три, реже в четыре этажа. В середине располагаются собственно эмбриональные клетки, из которых впоследствии разовьется зародыш. Очень редко возникает несколько зародышей, из которых полного развития достигает обычно только один, имеющий более длинный суспензор.
Головчатотисс Форчуна - обычно деревья высотой до 12 м, а иногда и кустарники. У деревьев кора красновато-коричневая, продольно слущивающаяся, листья длиной 5-8 см, шириной 4-5 мм, с постепенно заостряющейся верхушкой. Растет этот вид только в горах Центрального и Южного Китая.
Головчатотисс Харрингтона - деревья высотой до 15 м или кустарники. Кора у них, в отличие от предыдущего вида, серо-бурая. Листья короче, длиной 1,8-4,5 см и шириной 2-3,5 мм, к верхушке внезапно заостренные, на боковых побегах двурядно расположенные, часто кверху приподнятые (рис. 223). Этот вид головчатотисса распространен в Центральном Китае и в Японии. Растет обычно на высотах 700-2600 м, под пологом лиственных и хвойных лесов, в областях, богатых осадками.

Рис. 223. Головчатотисс Харрингтона (Cephalotaxus harringtonia): 1 - побег со взрослыми листьями; 2 - лист с нижней стороны; 3 - ветвь с микростробилами; 4 - часть побега с семенами; 5 - побег ювенильной формы
Головчатотиссы Форчуна и Харрингтона широко используются в качестве декоративных растений. Из них второй интродуцирован в Европу уже в 1829 г. В СССР культивируется почти исключительно в южных ботанических садах (Крым, Кавказ), где дает зрелые семена.
Отметим, что в культуре встречается специально выведенная пирамидальная или колонновидная садовая форма - головчатотисс Харрингтона, островерхая форма (С. harringtonia f. fastigiata), отличающаяся характером ветвления и листорасположения: боковые побеги направлены вверх и прижаты к главному стеблю, все листья, в том числе и на боковых побегах, спирально расположены. По-видимому, эта культигенная форма получена путем черенкования молодых растений.
При посеве семян головчатотисса Харрингтона молодые растения в течение первых 3-4 лет имеют такое же спиральное листорасполо-жение, как и у островерхой формы. Иногда в кроне типичного головчатотисса Харрингтона вместо побегов с гребенчато-двурядно расположенными листьями появляются отдельные боковые побеги со спиральным листорасположением.
В Японии из семян головчатотисса получают воскоподобное вещество, ядовитое, пригодное для технических целей (например, для свечного производства), а масло семян используется для приготовления красок и лаков.
Семейство тиссовые (Тахасеае) (О. А. Муравьева, В. С. Борхвардт)
Тиссовые, в отличие от араукариевых, распространены в северном полушарии (карта 35). Исключение составляют род австротаксус (Austrotaxus) с единственным видом в Новой Каледонии, а также тисс целебесский (Taxus celebica), переходящий экватор и встречающийся до 5° 30' ю. ш. в Индонезии на островах Суматра и Сулавеси. В северном же полушарии ареал тиссовых обширный и, как у многих других древних групп растений, разорванный. Они встречаются и в Северной Америке, и в Европе, и в Азии. Древность этого семейства, тесно связанного родством с головчатотиссовыми и также с подокарповыми, подтверждается и палеоботаническими данными. Бесспорные представители тиссовых известны уже с юры. Таковы находки тисса юрского (Taxus jurassica) в Йоркшире (Великобритания) и палеотаксуса оживающего (Palaeotaxus rediviva) в Швеции.
В семействе тиссовых 5 родов. Это тисс (Taxus), торрея (Тоггеа), псевдотаксус, или лжетисс (Pseudotaxus), австротаксус (Austrotaxus) и аментотаксус (Amentotaxus).
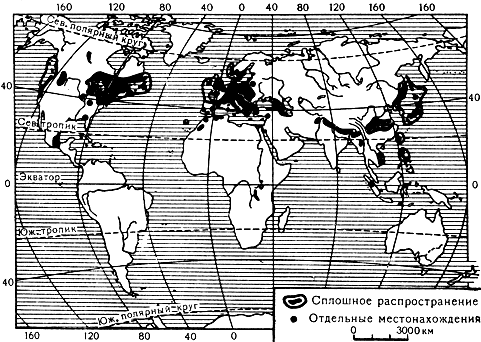
Карта 35. Ареал рода тисс
Тиссовые - вечнозеленые деревья или кустарники. В строении древесины с ее более или менее отчетливо выраженными годичными слоями прироста характерно наличие в трахеидах вторичных спиральных утолщений, которых нет только у австротаксуса, а также отсутствие в древесине смоляных каналов. Что касается древесинной паренхимы, то между родами семейства тиссовых наблюдаются определенные различия - то она скудная (тисс), то более или менее обильная (аментотаксус, австротаксус), то, наконец, полностью отсутствует (псевдотаксус).
Листорасположение очередное, но на боковых горизонтально отходящих ветвях оно почти супротивное, при этом листья благодаря изгибанию черешков обычно располагаются более или менее двурядно.
Листья тиссовых ланцетовидные или линейные (даже узко линейные), с низбегающим основанием, иногда на коротких (1-2 мм) черешках. На верхней стороне листа посредине тянется продольное углубление, на дне которого чуть выступает средняя жилка, снизу же между хорошо выраженной средней жилкой и зелеными, лишенными устьиц полосками по краям листа проходят две светлые устьичные полоски, то узкие, то более широкие.
Тиссовые двудомны, редко однодомны. Последнее отмечено у культивируемых растений торреи, иногда также у тисса.
Микростробилы у большинства тиссовых одиночные, но встречаются также их сережковидные, колосовидные или шаровидные (головчатые) собрания, расположенные в пазухах листьев. Микроспорофиллы имеют различное строение: то они щитковидные - состоят из ножки и щитка, к которому снизу при креплены 5-9 микроспорангиев, срастающихся между собой и с ножкой, как у тисса, то плоские дорсивентральные с четырьмя свисающими свободными микроспорангиями, как у торреи, то из одной ножки, от верхушки которой отходят 4-6 также свободных микроспорангиев, как у псевдотаксуса. Микроспоры у тиссовых без воздушных мешков (табл. 64).
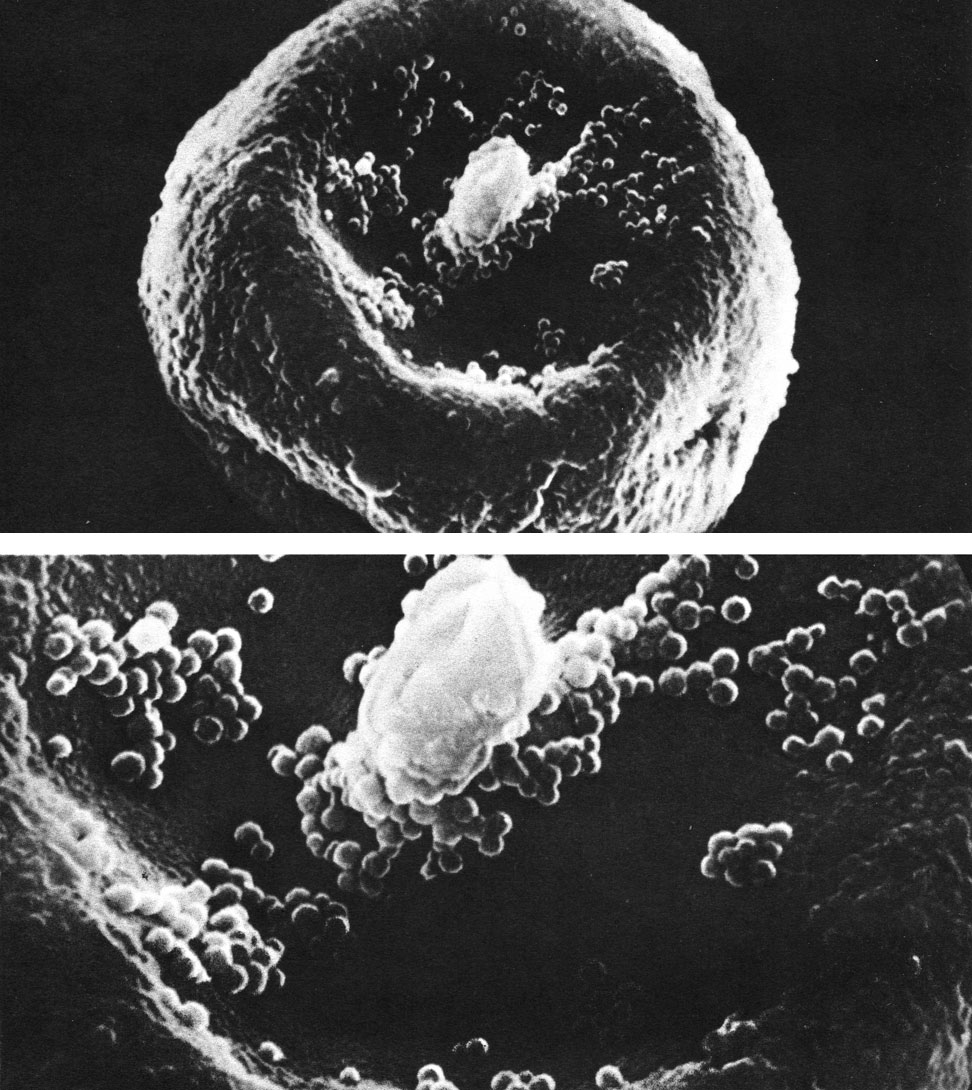
Таблица 64. Пыльцевое зерно тисса ягодного (Taxus baccata) под сканирующим электронным микроскопом (вверху - увел, около 5000, внизу - увел, около 13000)
Мегастробилы объединены в собрания, имеющие вид мелкой шишки с супротивно расположенными парами мегастробилов. Обычно же эти собрания сильно редуцированы и сведены к одиночным мегастробилам. По своему строению мегастробилы тиссовых более всего похожи на мегастробилы головчатотиссовых и некоторых подокарповых. Они состоят всего из одного прямого семязачатка, позднее семени, свободно (без срастания) окруженного мясистой бокальчатой кровелькой, или как ее называют, ариллусом, красного, желтого или белого цвета. У торреи семя костянковидное. Оливковая с пурпурными полосками кровелька покрывает семя полностью (до верхушки) и срастается с ним.
От характеристики семени вернемся к моменту опыления и к своеобразным, свойственным тиссовым эмбриологическим процессам на примере тисса, отчасти и торреи.
Семязачаток у тисса возникает поздней осенью перед годом оплодотворения и перезимовывает в стадии образования мегаспороцитов - материнских клеток мегаспор. У тисса в одном нуцеллусе бывает от 2 до 4 женских гаметофитов, возникающих из мегаспор одной и той же тетрады, однако не каждый из них образует архегонии.
Архегонии у тисса, а особенно у торреи, очень мелкие, окруженные отчетливо выраженным кроющим слоем из прилегающих клеток гаметофита. Укажем попутно, что именно у тисса ягодного (Taxus baccata) в оболочке яйцеклетки, прилегающей к клеткам кроющего слоя, впервые (в 1883 г.) русским ботаником И. Н. Горожанкиным были открыты плазмодесмы, пронизывающие пленки многочисленных здесь пор. Число архегониев у тиссовых различно: у тисса от 5 до 8, иногда до 11 и даже до 17, у торреи калифорнийской (Torreya californica) их 2-4, а у торреи тиссолистной (Т. taxifolia) всего один и лишь очень редко два. Изредка (у тисса и торреи) наблюдались архегониальные комплексы из 2 архегониев, заключенных в общий кроющий слой.
Ко времени рассеивания микроспор они у тисса одноклеточные, а у торреи двуклеточные. Проталлиальные клетки отсутствуют. Дальнейшее развитие мужского гаметофита осуществляется уже после опыления, когда пыльцевое зерно попадает на верхушку нуцеллуса. Именно здесь у тисса ядро пыльцевого зерна (в данном случае фактически микроспоры) делится, в результате чего в нем возникает маленькая генеративная клетка. При прорастании пыльцевой трубки генеративная клетка делится, образуя стенную клетку, или клетку-ножку, у которой скоро исчезает оболочка, и более крупную сперматогенную клетку. Позднее в пыльцевую трубку переходит все содержимое пыльцевого зерна: ядро пыльцевой трубки, ядро стенной клетки и сперматогенная клетка, которая лишь перед самым оплодотворением делится на 2 неравной величины спермия. Оплодотворение у тисса происходит в конце мая - начале июня. В целом развитие мужского гаметофита, благодаря перерыву на зимний период, продолжается очень долго, около шести с половиной месяцев. Напротив, от опыления (в марте) до оплодотворения проходит всего около месяца, после чего семена созревают за 6 недель.
В роде около 8 видов, встречающихся, как отмечалось, почти исключительно в северном полушарии (карта 35).
Тиссы - главным образом деревья, но иногда и кустарники. Ствол молодого дерева гладкий. Для старых тиссов характерна глубокая продольная бороздчатость. Кора красноватая или красновато-коричневая.
Листья на побегах, направленных вверх, расположены спирально, на горизонтальных же - двурядные, почти гребенчатые, линейные, иногда слегка серповидно согнутые. Сверху лист с выдающейся продольной жилкой, снизу - с двумя желтовато-зелеными или се-роватыми устьичными полосками. Характерной особенностью рода является полное отсутствие в листе смоляных каналов.
Микростробилы почти шаровидные (с перекрестнопарными пленчатыми чешуями при основании), на коротких ножках, покрытых очень мелкими чешуйками. Каждый стробил несет от 6 до 14 щитковидных микроспорофиллов, а каждый микроспорофилл - 5-9 микроспорангиев (рис. 224).
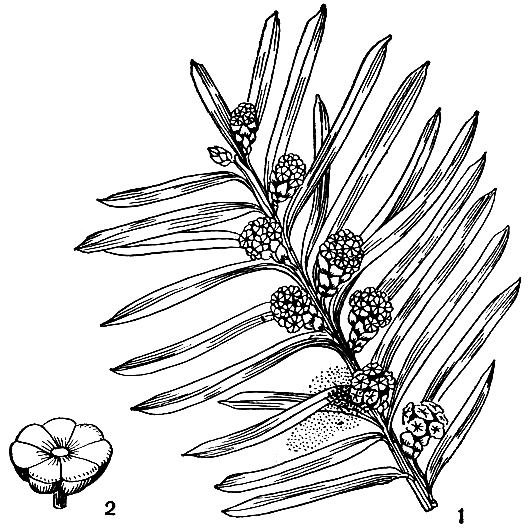
Рис. 224. Тисс ягодный (Taxus baccata): 1 - ветвь с микростробилами; 2 - микроспорофилл
Мегастробилы у тиссов одиночные на концах коротких сильно редуцированных пазушных побегов. Ось побега покрыта очень мелкими спирально расположенными чешуйками. Стробил несет 1, реже 2, совсем изредка 3 семязачатка.
Зрелое семя обычно овально-яйцевидное, длиной 5-8 мм, шириной 4-5 мм, окружено бокальчатой, мясистой, обычно красной, реже желтой кровелькой. Кровелька обнимает семя до верхушки или до половины, но с ним не срастается (табл. 63).
Наиболее известным и шире других распространенным представителем рода является тисс ягодный, или обыкновенный (Taxus baccata). Называют его нередко и европейским, поскольку он произрастает почти по всей Западной Европе. Ареал тисса ягодного охватывает, кроме того, районы Западной Белоруссии (Беловежская пуща), Западной Украины (Буковина), Южного Крыма, Кавказа, а также Азорские острова, горы Алжира, Малой Азии и Сирии. В народе широко бытует еще одно название - "негной-дерево", отражающее очень важные для хозяйственного использования свойства древесины тисса. Она твердая и тяжелая, почти не поддается гниению. Действительно, тисс исключительно устойчив по отношению к грибным заболеваниям и поражениям насекомыми, хотя в определенных условиях может от них страдать. Самым опасным грибом для него является, по-видимому, полипорус серно-желтый (Polyporus sulphureus), который вызывает заболевание, известное под названием "сердцевинная гниль древесины". Иногда на тиссе наблюдаются наплывы, густо покрытые очень короткими побегами с бледной хвоей, так называемые "ведьмины метлы".
Продолжительность жизни тисса ягодного очень велика - до 1500 лет, а иногда, по-видимому, и до 3-4 тыс. лет.
Годичный прирост составляет 2-3 см, поэтому деревья не дорастают до большой высоты, однако известны отдельные крупные экземпляры, по которым можно судить о долголетии тисса. Так, в Англии, в Дербишире известен тисс высотой 15 м, возраст которого оценивается в 2000 лет. Тисс в графстве Суррей уже в 1133 г. был известен как очень старый экземпляр. Самым высоким деревом со стволом высотой 32,5 м и диаметром 1,5 м, по всей вероятности, является тисс, растущий в Закавказье (Аджария). Там же имеются старые многовершинные экземпляры с диаметром ствола у основания до 2,5 м.
Ствол тисса ягодного покрыт красновато- коричневой или красновато-бурой корой. Она гладкая, позднее трещиноватая, продольно отслаивающаяся. Густая крона имеет яйцевидно-коническую, реже пирамидальную форму.
На нижней стороне облиственных побегов, по одному в пазухах вегетативных листьев, расположены многочисленные шаровидные микростробилы. В пазухах листьев на коротких боковых веточках сидят и мегастробилы с одним, реже с двумя семязачатками. Семена до самой верхушки окружены ярко-красной мясистой кровелькой, что делает их очень похожими на ягоды, с чем и связан видовой эпитет этого растения (табл. 63).
Тисс ягодный требователен к влажности воздуха и почвы. Это и самое теневыносливое из всех хвойных пород растение, почему его часто можно встретить растущим совместно с самым теневыносливым цветковым растением - самшитом. В то же время тисс растет и на открытых местах. Обычно сам он чистых насаждений не образует, в молодости лучше развивается под пологом бука, граба и пихты. Семена тисс дает ежегодно до глубокой старости, начиная с 25-30-летнего возраста.
Молодые побеги, кора и листья содержат таксин - алкалоид, ядовитый для человека, а также для некоторых домашних животных, например лошадей и коров. Но другие животные, например зайцы, олени, поедают тисс охотно и без вреда для себя. Семена тисса тоже содержат таксин, но в мясистой сладковатой кровельке ядовитых веществ нет, поэтому птицы (черный дрозд), а из мелких зверей куница поедают семена и частично разносят их, способствуя распространению тисса. Естественное возобновление под материнским пологом обычно не наблюдается, всходы и подрост встречаются в стороне от взрослых деревьев, иногда на значительном расстоянии. Тисс обладает высокой побегопроизводительной способностью, что очень важно для культурных насаждений; на пнях дает обильную поросль. Отмечены случаи укоренения нижних ветвей при соприкосновении с землей.
Корневая система тисса хорошо развита, пластична, обладает эндотрофной микоризой, благодаря чему он может приспосабливаться к разным условиям, встречается как на рыхлых свежих, так и на плотных каменистых почвах.
Растет тисс обычно от уровня моря до высоты 1500 м, но чаще в поясе от 500 до 1200 м. Изредка он достигает верхней границы леса, где приобретает вид приземистых кустарников, зимует под снегом, но при этом не плодоносит.
Чаще всего тисс ягодный встречается во втором ярусе буково-грабовых или смешанных лесов из бука, пихты кавказской и ели восточной, растет единичными экземплярами или небольшими группами. На Кавказе сохранились и такие участки, где тисс имеет лесообразующее значение. Такова, например, заповедная Хостинская тиссо-самшитовая роща, где тисс, являясь господствующим растением, занимает площадь около 70 га. Самое крупное естественное место произрастания тисса находится в Восточной Грузии, по ущелью Бацара, в верховьях реки Алазани. Эта роща занимает площадь около 700 га. По свидетельству крупнейшего знатока лесов Кавказа А. Г. Долуханова, эта тиссовая роща "по размерам, по числу взрослых деревьев, по величественности древостоя, по мощности роста и другим признакам является не только лучшей на Кавказе, но и лучшей на всем земном шаре". Общая площадь, занимаемая тиссом на Кавказе, составляет более 1500 га.
Когда мы говорим о древесине тисса, то, как правило, прибегаем к народным его названиям "негной-дерево", о смысле которого мы уже рассказывали, или "красное дерево", поскольку древесина тисса действительно красная. Красный цвет древесины под действием воды становится пунцово-фиолетовым, а от долгого пребывания в воде - почти черным.
Древесина тисса - прекрасный материал для строительных, столярных, токарных работ, отлично полируется, что важно в мебельном производстве. Из нее уже в глубокой древности, за 3000 лет до нашей эры, сооружали погребальные саркофаги, позднее делали луки и разнообразные хозяйственные изделия. Широкое использование тисса привело к его массовому уничтожению.
Являясь довольно выносливым вечнозеленым растением, тисс ягодный широко используется и в зеленом строительстве, в частности, в южных и юго-западных районах СССР. Он ценится за темную зелень листвы, за то, что безболезненно переносит стрижку и формирование кроны, которой придают художественные формы шаров, пирамид и даже подобия животных. Поскольку тисс растет очень медленно, то эти формы сохраняются долго. Во время созревания семян он очень эффектен из-за яркой окраски кровельки. Тисс относительно дымоустойчив. Наконец, он хорошо переносит пересадку во взрослом состоянии. Посеянные семена всходят через 1-2 года, иногда через 3 года, сохраняя всхожесть в течение 4 лет. Можно разводить тисс не только семенами, но и черенками и отводками.
Тисс ягодный, как ценное растение, вводился в культуру с древних времен. К настоящему времени описано до 50 его садовых форм. Многие из них отличаются высокой декоративностью, красивой формой роста и окраской листвы. Однако то обстоятельство, что листья тисса ядовиты, что домашние животные, поедавшие листья и молодую поросль, заболевали, вызывало недоброжелательное отношение к нему со стороны местного населения, и люди зачастую сознательно вырубали эти замечательные растения.
Второй известный представитель рода во флоре Евразии - тисс остроконечный (Taxus cuspidata), как показывает название, отличается от своего европейского собрата прежде всего листьями, оканчивающимися не коротким шипиком, как у последнего, а игольчатым остроконечием. Впрочем, более существенным является отличие в строении семени. Оливково-коричневые семена здесь всего наполовину, а то и менее погружены в сочную кровельку. В остальном сходство между этими тиссами настолько велико, что ботаники начала прошлого столетия считали их одним видом. Но если тисс ягодный является в основном западноевропейским видом, тисс остроконечный распространен на противоположной, восточной окраине Евразиатского материка. Его ареал охватывает здесь южную часть советского Дальнего Востока, включая горную систему Сихотэ-Алиня, остров Сахалин и Южные Курилы, а также территории соседних стран - Кореи, Китая (включая Тайвань), Японии.
Дальневосточные ботаники характеризуют тисс остроконечный как одно из самых оригинальных и редко встречающихся хвойных рас-тений крайнего востока нашей страны. Он обитает здесь на высоте от 100 до 900 м над уровнем моря, в богатых, реликтовых (по С. И. Коржинскому) хвойно-широколиственных лесах. Еще издали вы узнаете старый тисс среди стволов разнообразных деревьев по толстому (диаметром до 1 м и более), часто, как у тисса ягодного, глубокобороздчатому стволу. Узнается он и по гладкой, очень тонкой, красной или красно-бурой коре, отличающей его от всех деревьев-соседей. Подойдя ближе, можно рассмотреть и крону тисса, низко посаженную и широкую, образованную горизонтально отклоненными ветвями, одетыми темно-зелеными листьями. Теневыносливый тисс остроконечный, достигающий высоты обычно 10- 15, редко 20 м, мирится с сильным затенением кронами лесных гигантов дальневосточных лесов. На фоне темно-зеленых блестящих листьев красиво выделяются "полуодетые" красной кровелькой семена.
В пределах всего ареала тисс остроконечный, как правило, не образует самостоятельных, тем более чистых насаждений (тиссовых лесов), встречаясь или отдельными деревьями, или лишь небольшими группами. Возобновление тисса встречается исключительно редко и главным образом в виде "карликовых" одно-двухгодичных экземпляров (тисс растет медленно и даже в возрасте 30 лет достигает всего 1-1,2 м высоты). Это кажется парадоксальным, поскольку тисс остроконечный хорошо плодоносит и нормально размножается семенами, имеющими высокую всхожесть. Объясняется почти полное отсутствие возобновления тем, что птицы и мелкие грызуны в массе поедают его семена и, с другой стороны, копытные животные обгрызают верхушки подроста. В этом отношении исключение составляют некоторые острова у берегов Приморского края, особенно остров Петрова, расположенный в Японском море в 600-700 м от берега, в районе бухты Преображения. Здесь отсутствуют копытные, меньше птиц и грызунов и, как следствие этого, тисс обилен, возобновляется отлично, образует почти чистые насаждения; одно из них на острове Петрова занимает площадь около 1 га.
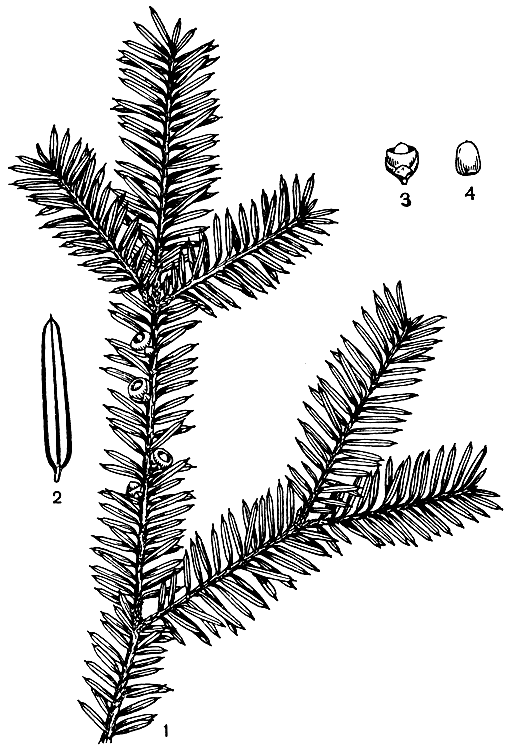
Рис. 225. Псевдотаксус Цзяня (Pseudotaxus chienii): 1 - ветвь с мегастробилами; 2 - лист; 3 - семя с ариллусом и стерильными чешуями; 4 - зрелое семя без ариллуса
На ряде островов тисс вместе с древесными растениями из цветковых приобретает стелющуюся форму, образуя непроходимые заросли. Впрочем, кустарниковая форма тисса остроконечного наиболее характерна для северозападной части ареала, где она приурочена к более открытым, освещенным местам - редкостойным лесам, безлесным склонам гор. Кустарниковый тисс развивает простертые, прижатые к земле ветви длиной до 3 м (диаметр всего куста до 5 м). Нижние ветви его часто укореняются, давая начало дочерним кустам.
Семена тисса остроконечного, похожие на кедровые орехи, употребляет в пищу местное население. Этот дальневосточный тисс, как и тисс ягодный, дает ценную древесину. Однако, учитывая редкость этого замечательного растения и сокращение его природных запасов, оно объявлено объектом охраны в "Красной книге". В Европе тисс остроконечный введен в культуру как декоративное растение; хорошо растет в Англии, Франции и других странах Европы, в восточных штатах США. В СССР растет и дает семена в ботанических садах Ленинграда, Москвы и других городов.
Тисс канадский (Т. canadensis) распространен от восточной провинции Ньюфаундленд в Канаде, через область Великих озер до штата Вирджиния в США. Это однодомный низкий раскидистый кустарник высотой около 1-2 м, с отклоненными, простертыми, приподнятыми вверх ветвями. Побеги у него короткие, густооблиственные, почки яйцевидные или эллипсоидальные, мелкие, желто-зеленого цвета. Почечные чешуи ланцетовидные.
Тисс канадский - растение холодостойкое. Он выносит морозы до -30, -35°С. Встречается преимущественно в низменных влажных местах, иногда по скалистым горным склонам, до 740 м над уровнем моря. Растет в подлеске хвойных лесов, иногда в хвойно-широколиственных лесах. Введен в культуру с 1800 г., изредка разводится в Англии, Франции, Италии. В СССР в ботанических садах Ленинграда и Москвы достаточно зимостоек и дает семена (табл. 63).
Тисс коротколистный (Т. brevifolia) распространен в западной части Северной Америки, от Британской Колумбии в Канаде до Калифорнии в США. Растет по берегам рек и приозерным низменностям, на невысоких горных склонах и в глубоких ущельях, на богатых, хорошо дренированных почвах. На севере спускается до уровня океана, на юге растет в горах, на высоте 1500-2400 м. Чистых насаждений этот тисс не образует, встречается единично или группами во втором ярусе лесов.
Деревья этого вида имеют много общего с тиссами ягодным и остроконечным. Высотой от 6 до 15 м (редко до 25) и со стволом в диаметре до 30-40 см, они несут ширококоническую крону, обычно с горизонтально простертыми или восходящими ветвями и слегка поникающими молодыми веточками. Кора красновато-коричневая. В северной части ареала встречается в виде распростертого кустарника.
Тисс коротколистный интродуцирован в Европу с 1854 г., культивируется как декоративное растение в садах и парках Англии, Франции, Италии. Растет в Ленинграде.
Род псевдотаксус монотипный. Обычно это двудомные кустарники высотой 2-4 м, с мутовчатыми или супротивными ветвями, иногда небольшие деревья (рис. 225).
Листья длиной до 2,5 см и шириной 2-3 мм, густо спирально расположены на боковых ветвях, двурядные, иногда почти супротивные, узколинейные, сверху зеленые и несколько выпуклые. Верхушка листа остроконечная, основание слегка асимметричное, внезапна суженное в короткий черешок. Устьица на нижней стороне, иногда и на верхней. Гиподермы и смоляных каналов нет. В отличие от тисса нижняя эпидерма гладкая. Микростробилы на очень коротких ножках в пазухах зеленых листьев. Кроме микроспорофиллов они несут по 2-3 стерильные чешуйки, чередующиеся с микроспорофиллами. Мегастробилы длиной до 7 мм, одиночные, пазушные. Кровелька появляется позднее интегумента, окружает семязачаток лишь ко времени опыления, но с интегументом не срастается; нуцеллус также свободен от интегумента. Зрелое семя длиной 6 мм окружено мясистой, колокольчатой кровелькой белого цвета, не срастающейся с семенем в его верхней части.
Псевдотаксус является эндемиком Восточного Китая, растет там в горных лесах на высоте до 1000 м над уровнем моря.?
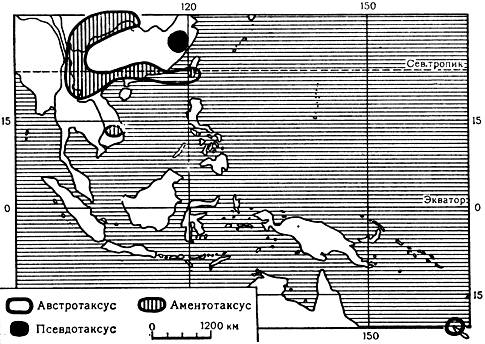
Карта 36. Ареалы родов австротаксус, аментотаксус и псевдотаксус
Род торрея (Torreya). Сюда относятся вечнозеленые двудомные, реже однодомные деревья, с супротивными или даже мутовчатыми молодыми побегами. Почки покрыты многочисленными супротивными чешуями.
Листья очередные, реже супротивные, на боковых побегах двурядно гребенчато расположенные, длиной до 4 и даже 6 см, линейные или линейно-ланцетовидные, жесткие, к верхушке постепенно заостренные в колючку. На их нижней поверхности по обе стороны от жилки проходят узкие беловатые, с возрастом рыжевато-буроватые устьичные полоски. При растирании листья некоторых видов распространяют сильный неприятный запах. Эпидерма листа (как верхняя, так и нижняя) состоит из клеток столбчатой формы с очень толстой одеревеневшей оболочкой и узкой, иногда в виде щели полостью; снаружи эпидерма покрыта довольно толстым слоем кутикулы. Мезофилл листа состоит из двух, реже трех слоев палисадной паренхимы, а также из губчатой паренхимы, клетки которой вытянуты поперек. По обе стороны жилки на нижней поверхности листа находятся углубления, в которых погружены устьица; над замыкающими клетками устьиц, покрывая их, развиты длинные папиллы - выросты побочных клеток, часто заканчивающиеся двулопастной верхушкой. Их наличие является уникальной особенностью рода торрея. Довольно длинные, но совершенно цельные папиллы имеются только у сросшихся попарно листьев рода сциадопитис (Sciadopitys).
Микростробилы, по одному в пазухах вегетативных листьев, почти шаровидные или эллипсоидальные, длиной до 8 мм, окружены 4 па-рами стерильных чешуй. Микроспорофиллы по 4 собраны в 6-8 мутовок; каждый микроспорофилл с короткой ножкой и расширенной дорсивентральной верхушкой несет 4 свободных, свисающих микроспорангиев, раскрывающихся продольной щелью.
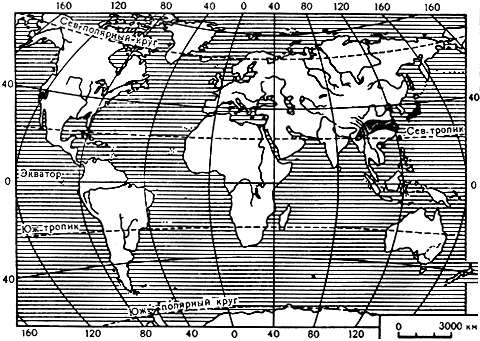
Карта 37. Ареал рода торрея
Мегастробилы, как и микростробилы, располагаются в пазухах зеленых листьев на побегах последнего года (рис. 226). Они состоят из короткой оси, несущей пару семязачатков (реже 2-3 пары). Каждый семязачаток находится в пазухе кроющего листа и одет двумя парами стерильных чешуй. Базальная часть, молодого семязачатка окружена кровелькой в виде валика. В дальнейшем благодаря вставочному росту нижней части кровельки ее: верхняя свободная часть (архисперм) выносится кверху, а разросшаяся нижняя, называемая гипоспермом, облекает семя и срастается с интегументом. Из двух семязачатков стробила обычно развивается и дает семя только один, редко 2 и даже 3.
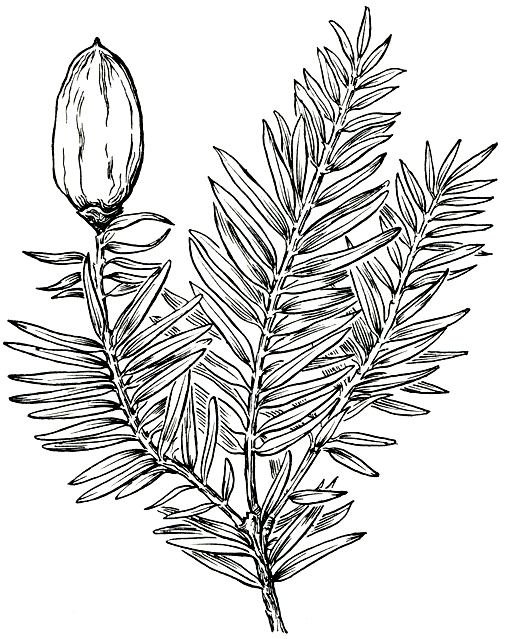
Рис. 226. Торрея орехоносная (Torreya nucifera), ветвь с мегастробилом
Зрелое семя с кровелькой, овальное или широкоовальное, длиной до 2,5 см, с наружным (из кровельки) мясистым и внутренним (из интегумента) деревянистым слоем, зеленовато-оливковое с пурпурными полосками или пурпурное. Само семя содержит руминированный (морщинистый) эндосперм и очень мелкий зародыш с двумя слабо развитыми семядолями. В отличие от тисса семена здесь прорастают не надземно, а подземно.
Ареал рода торрея, насчитывающего 6 видов, разорванный. Две разъединенные части ареала в Северной Америке: одна на западе, в Калифорнии, вдоль западных склонов гор Сьерра-Невада, другая - в северо-западной части штата Флорида и юго-западной части штата Джорджия. Еще два отделенных друг от друга участка ареала торреи находятся в Восточной Азии: один из них в Японии, другой - в Китае и Бирме.
Торрея калифорнийская, или мускатная (Torreya californica), растет по склонам Сьерра-Невады, поднимаясь до 1500 м над уровнем моря, иногда встречается и по долинам рек. Деревья относительно небольшой высоты (10- 15 м, редко до 35 м), диаметр ствола 1 -1,2 м, иногда встречается и в виде кустарника. Простертые, несколько поникающие на концах ветви этой торреи образуют пирамидальную, позднее округлую крону. Кора серовато-коричневая.
Древесина, семена и листья торреи калифорнийской обладают резким запахом.
Торрея калифорнийская интродуцирована в Европу в прошлом столетии. Растет на Южном берегу Крыма (Никитский ботанический сад, Артек), изредка на Черноморском побережье Кавказа.
Торрея крупная (Т. grandis) произрастает в горах Северо-Восточного и Центрального Китая в виде деревьев высотой 17-25 м, с серовато-коричневой корой, а иногда в форме кустарника.
В культуре встречается очень редко как в Европе, так и в Северной Америке. В настоящее время торрея крупная лишь изредка встречается в дендрариях и ботанических садах на Черноморском побережье Кавказа.
Торрея орехоносная (Т. nucifera) встречается на высоте 500-1800 м над уровнем моря, чаще всего во втором ярусе хвойных и широко-лиственных лесов, в горах Южной и Центральной Японии. Это деревья высотой 10-25 (до 30) м, с плотной пирамидальной кроной и простертыми ветвями.
Семена съедобные, их используют в пищу и для приготовления пищевого и технического масла, на лаки и краски. Древесину используют в строительстве и столярном производстве.
Этот вид торреи, интродуцированный в Европу более ста лет назад, ныне успешно растет и дает семена. Он культивируется на Черноморском побережье Кавказа, иногда дает здесь семена. В Батумском ботаническом саду 4-летний экземпляр этой торреи достигал высоты 11 м и диаметра около 25 см.
Размножается торрея орехоносная вегетативно, ее можно прививать, как и другие виды торреи, на тиссе ягодном, а также размножать семенами. Семена, прорастающие обычно только на второй год, рекомендуется стратифицировать. Как декоративное растение очень эффектно благодаря красивой форме кроны, блестящей темно-зеленой листве. Из всех торрей этот вид - наиболее холодостойкий.
Торрея тиссолистная (Т. taxifolia) распространена на северо-западе штата Флорида в США на известняковых обрывах гор. Это деревья с пирамидальной кроной, высотой до 12-15 м, со стволом в диаметре до 60 см. Кора их неправильно трещиноватая, чешуйчатая, коричневая с оранжевым оттенком. Темно-зеленые листья торреи тиссолистной, как явствует из названия, напоминают листья тисса, но крупнее их (длиной до 3 и даже 4 см).
Широкоэллипсоидальные или обратнояйцевидные, темно-пурпурные семена этого вида достигают длины 3-4 см.
При растирании листья торреи тиссолистной и ее семена распространяют резкий запах.
В Европу это растение интродуцировано уже в 1838 г., но встречается редко. В Батумском ботаническом саду имеется один не дающий семян экземпляр.
Название этого рода происходит от латинских слов auster - южный и taxus - тисс. В нем всего один вид - австротаксус колосистый (A. spicata), распространенный в южном полушарии, во влажных горных лесах северной половины острова Новая Каледония, преимущественно на сланцевых и гнейсовых скалах на высоте 400-1600 м над уровнем моря. В культуре этот вид не встречается.
Это вечнозеленые, довольно высокие (15-25 м) деревья, с сероватой, морщинистой корой и густооблиственной ветвистой кроной. Особое внимание привлекают листья деревьев - длинные, узколинейно-ланцетовидные, с загнутыми вниз краями. Длина их от 10 до 15 см при ширине до 1 см.
Микростробилы составляют прямостоячие, колосовидные собрания; ось стробилов несет 12-15 чешуевидных кроющих листьев, а в их пазухах располагаются по одному элементарному сильно редуцированному стробилу, состоящему из 1-3 (до 5) микроспорофиллов; каждый микроспорофилл состоит из короткой ножки, несущей на верхушке 3, реже 2 или 4 микроспорангия.
Мегастробилы верхушечные, на концах коротких веточек, густо покрыты спирально расположенными чешуйками. Семязачаток прямой, вначале лишь у основания окружен кровелькой, которая позднее разрастается до верхушки семязачатка, но не срастается с ним. Зрелое семя полностью заключено в мясистую кровельку.
Впервые это растение было описано в 1922 г. Долгое время его систематическое положение оставалось неопределенным и даже спорным, так как по морфологическим, анатомическим и эмбриологическим признакам он занимает промежуточное положение между семействами подокарповых и тиссовых. По внешнему облику, по характеру листьев, по наличию сложных колосовидно собранных микростробилов, по отсутствию типичных спиральных утолщений у трахеид вторичной ксилемы, наконец, по характеру развития предзародыша австротаксус имеет большое сходство с некоторыми видами подокарповых. Казалось бы, и произрастание его в южном полушарии, где распространены подокарповые и отсутствуют представители тиссовых, тоже свидетельствует о близости этого рода к семейству подокарповых. Однако у австротаксуса колосистого есть ряд важных признаков, общих с семейством тиссовых. Так, семязачатки у него прямые, одиночные, верхушечные, окруженные симметричной кровелькой; число микроспорангиев больше, чем у подокарповых, микроспоры без воздушных мешков. В то же время авсгротаксус отличается от тисса формой и величиной архегониев, очень длинными первичными суспензорами и некоторыми другими признаками.
Вот почему в 1938 г. японский ученый Т. Накаи попытался разрешить споры о систематическом положении австротаксуса выделением его в самостоятельное семейство, что и нам представляется справедливым, однако и теперь неясность положения сохраняется.
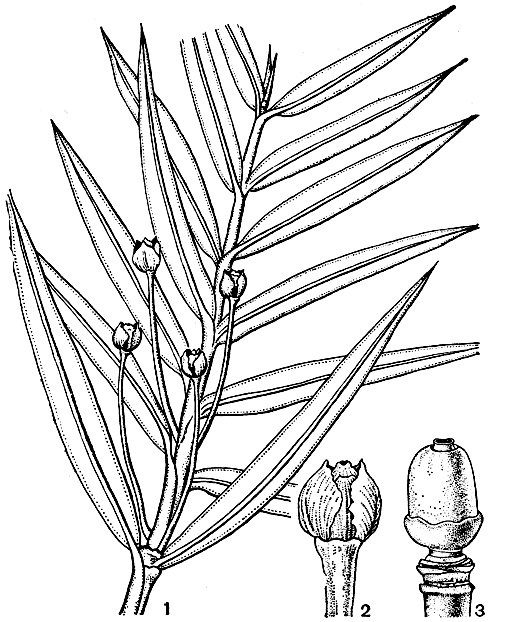
Рис. 227. Аментотаксус серебристый (Amentotaxus argotaenia): 1 -ветвь с мегастробилами; 2 - мегастробил; 3 - семязачаток с удаленными чешуями
Это родовое название происходит от латинских слов amentum - сережка и taxus - тисс, и дано оно роду потому, что самой характерной особенностью рода является наличие сережковидных свисающих микростробилов. Род включает 4 вида. Все они распространены в юго-восточной части Китая, включая остров Тайвань, на западе Китая и в северной части Вьетнама, обычно в ущельях, на склонах гор и по берегам горных речек на высоте от 300 до 1600 м в тенистых местах.
Это небольшие вечнозеленые деревья (высотой до 10 м и диаметром ствола до 30 см), с раскидистой кроной или иногда кустарники высотой 2-3 м. Листья супротивные, на боковых ветвях двурядно расположенные в одной плоскости, линейные, прямые или слегка серповидно изогнутые, длиной 3,5-11 см и шириной до 1 см, кожистые, сверху темно-зеленые, с хорошо выраженной выступающей жилкой, на нижней поверхности с беловатыми устьичными полосками.
Микростробилы в сложных, сережковидных собраниях длиной до 3 см, повисающих, расположенных на концах вегетативных побегов по 3-4, редко по одному, а иногда и по пяти. Элементарные стробилы на очень коротких ножках, шаровидные, выходят по одному или по 2-3 из пазух сильно редуцированных чешуй. Микроспорофиллы почти дорсивентральные (как у торреи), несут по 4-5, реже по 3 свисающих микроспорангиев. Мегастробилы одиночные; (рис. 227), простые, на длинной ножке (до 2 см), выходящей из пазухи обыкновенных зеленых листьев.
Семязачаток у аментотаксуса прямой, почти шаровидный, окруженный 5-6 парами килеватых чешуй. Кровелька вначале окружает семязачаток лишь у основания. В дальнейшем благодаря вставочному росту базальной части кровельки ее верхняя свободная часть выносится кверху, а нижняя, разрастаясь, облекает семя и прирастает к интегументу, впоследствии отделяется от него. Ф. Оливер и Р. Флорин считают, что только верхняя, - свободная часть кровельки аментотаксуса соответствует всей кровельке тисса и псевдотаксуса, подобно тому как это имеет место у рода торрея, а нижняя, интеркалярная часть является новообразованием и составляет чашеобразное расширение оси стробила, охватывающее все семя и срастающееся с ним.
Представителей рода аментотаксус вначале относили к роду подокарп, поскольку известны были только их стерильные экземпляры. В 1903 г. Р. Пильгер отнес эти растения к роду головчатотисс (Cephalotaxus), а в 1916 г. выделил в особый род аментотаксус (Amentotaxus), характерной особенностью которого являются сложные, сережковидные микростробилы в сочетании с одиночными (простыми) мегастробилами. В 1926 г. Пильгер включил этот род в семейство головчатотиссовых. Однако аментотаксус своими репродуктивными органами очень сильно отличается от головчатотиссовых. Именно поэтому ряд ученых склоняется к выделению рода аментотаксус в самостоятельное семейство. К сожалению, виды этого интересного рода в культуре отсутствуют, нет их даже ни в одном из ботанических садов мира. Не удивительно поэтому, что аментотаксус относится к числу наименее изученных голосеменных.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'