Отдел псилотовидные (Psilotophyta) (А. Л. Тахтаджян)
Общая характеристика спорофита
Среди ныне живущих высших растений совершенно обособленное положение занимает маленький отдел псилотовидные (Psilotophyta), в который входит лишь один класс псилотопсиды (Psilotopsida), один порядок (Psilotales) и одно семейство псилотовые (Psilotaceae), состоящее из двух родов - псилота (Psilotum, рис. 74) и тмезиптериса (Tmesipteris, рис. 75). Первый из них, заключающий всего лишь два хорошо различающихся вида, довольно широко распространен в тропических и отчасти субтропических странах обоих полушарий и доходит на севере до Южной Испании, Южной Кореи, Южной Японии, Гавайских островов, юга США (Аризона, Техас, Луизиана, Флорида, побережье Южной Каролины) и Бермудских островов, а на юге достигает Новой Зеландии (острова Северного и островов Окленд). Второй же (тмезиптерис), состоящий из около 10 близких видов, распространен не столь широко. Его ареал охватывает Новые Гебриды, острова Самоа, ряд других островов Полинезии, остров Норфолк, Новую Каледонию, Австралию, Тасманию, Новую Зеландию, остров Стьюарт и острова Окленд, Чатем и Кермадек; на севере он достигает Филиппинских островов.
Псилотовые играют лишь очень скромную роль в современном растительном мире. Не имеют они также почти никакого практического значения в жизни человека, если не считать того, что псилот голый, или обыкновенный (Psilotum nudum, рис. 74), уже давно культивируется в Японии в качестве оригинального декоративного растения. И если мы уделяем в "Жизни растений" целую главу этим пасынкам растительного мира, то лишь потому, что они не только отличаются рядом интереснейших морфологических и биологических особенностей, но и представляют огромный интерес с точки зрения их происхождения. Как мы увидим дальше, в их строении сохранились чрезвычайно примитивные черты, которые свидетельствуют об их очень древнем происхождении. Такие растения неизменно вызывают большой интерес ботаников и всех интересующихся эволюционными аспектами ботаники, так как помогают лучше понять пути и направления эволюции растительного мира.
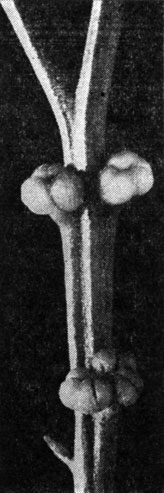
Рис. 74. Псилот голый (Psilotum nudum). Веточка с синангиями (увел.)
Как псилот, так и тмезиптерис растут на стволах древовидных папоротников, саговниковых или пальм, у основания стволов деревьев, на перегнойной почве или нередко в трещинах скал.
Растения обоих родов лишены корней. Их подземные органы представляют собой более или менее дихотомически разветвленные и довольно длинные (до 1 м и более) корневищеподобные образования, которые, в отличие от настоящих корневищ, лишены каких-либо чешуй, но покрыты многочисленными ризоидами. Морфологически эти подземные органы псилотовых вполне соответствуют ризомоидам древних, давно вымерших палеозойских риниофитов. Выходя на дневную поверхность, концы ветвей ризомоида развиваются в новые надземные стебли, чем достигается вегетативное размножение. Интересно, что не только поверхность эпидермальных клеток ризомоида, но и ризоиды покрыты кутикулой. В клетках его наружной коры часто содержатся гифы гриба, который проникает через ризоиды. Благодаря микоризным грибам псилотовые ведут частично сапрофитный образ жизни.
Интересно, что у псилота голого концы некоторых ризоидов разрастаются еще на ранней стадии развития в небольшую выводковую почку, или гемму (от лат. gemma - почка), которая способна дать начало новому ризомоиду, а следовательно, новому растению (табл. 15).

Таблица 15. Псилот голый (Psilotum nudum): вверху - общий вид куста в оранжерее Ботанического института Ан СССР в Ленинграде; внизу - веточки с синангиями
Отсутствие корней - одна из наиболее характерных особенностей псилотовых. Корней нет не только у взрослых растений, но зачаток корня (как и зачаток листа) отсутствует также на всех стадиях развития зародыша. Это дает основание предполагать, что отсутствие корня у псилотовых является скорее всего признаком первичным, а не вторичным, т. е. не является результатом редукции, как это нередко происходит у других растений. Отсутствие корня сближает псилотовые с риниофитами. С риниофитами их сближает и целый ряд других морфологических особенностей, доказывающих очень большую примитивность этих двух необычайно интересных родов.
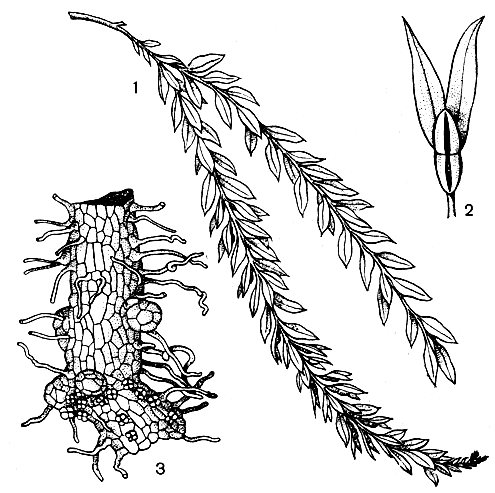
Рис. 75. Тмезиптерис (Tmesipteris tannensis): 1 - ветвь растения; 2 - веточка с синангием; 3 - гаметофит
Надземные оси (стебли) псилота повторно дихотомически (вильчато) ветвистые, причем ветвление здесь самого примитивного равнодихотомического типа. У тмезиптериса стебель чаще всего неразветвленный или же однажды (очень редко дважды) дихотомирован. У псилота стебли длиной от 10 до 100 см, а у тмезиптериса - от 5 до 25 (иногда до 40) см. Стебли эпифитных особей псилотовых обычно висячие, но у наземных они часто прямостоячие.
С эволюционной точки зрения большой интерес представляет вопрос о листовых органах псилотовых. На стеблях псилота, главным образом в верхней части, видны мелкие чешуевидные придатки, расположенные без определенного порядка. Они лишены как устьиц, так и жилок. Более листовидный характер носят эти придатки у тмезиптериса. В нижней части стебля они мелкие и чешуевидные, как у псилота, но выше становятся крупнее и представляют собой уплощенные латерально (в вертикальной плоскости) и нисходящие на стебель ланцетные или эллиптические пластинки, кончающиеся остроконечием. За исключением нижних чешуевидных придатков, они имеют хорошо выраженную жилку, представляющую собой одиночный проводящий пучок. В отличие от чешуевидных придатков эти пластинки снабжены с обеих сторон устьицами. Однако остроконечная верхушка этих пластинок, лишенная проводящего пучка и устьиц, сохраняет сходство с чешуевидными придатками. Как в свое время отметил Ф. Бауэр (1935), создается впечатление, что пластинчатая часть представляет собой новообразование, которое как бы приподняло вверх первоначальный чешуевидный придаток. Сами чешуевидные придатки возникли, по предположению Ф. Бауэра, в качестве выростов (эмергенцев) поверхностных тканей стебля, т. е. имеют энационное происхождение, а не теломное, как предполагают другие авторы. В этом отношении они сходны с плауновидными, у которых листовые органы также представляют собой выросты на стебле. Тенденция к образованию энациев имелась уже у риниофитов, в частности у некоторых видов псилофита (Psilophyton).
Если мы теперь обратимся к репродуктивным (спороносящим) частям спорофита, то и здесь найдем много интересных особенностей, в том числе определенные черты сходства с риниофитами. У псилотовых спорангии образуют синангии. У псилота каждый синангий состоит обычно из трех спорангиев, в то время как у тмезиптериса он двуспорангиевый. Синангии довольно крупные, диаметром 2-4 мм. Образование синангиев является, несомненно, признаком специализации и свидетельствует об определенной эволюционной подвинутости псилотовых.
Однако вспомним, что тенденция к образованию синангиев была выражена уже среди риниофитов, а у рода яравия (Yarravia) имелись настоящие синангии. Таким образом, в этом отношении псилотовые ушли не дальше некоторых специализированных риниофитов.
Несмотря на то что спорангии псилотовых срослись в синангии, они сохранили относительно примитивное строение. Каждый спорангий имеет толстую стенку, состоящую из 4-6 слоев клеток, и большое число спорогенных клеток. Но не все спорогенные клетки превращаются в спороциты (материнские клетки спор) - значительная их часть разрушается и превращается в питательную плазмодиальную жидкую массу, в которой развиваются функционирующие спороциты, а затем и споры.
Споры псилотовых обычно билатерально симметричные (монолетные), почковидные, с прозрачными оболочками и имеют сетчатую поверхность.
Каждый синангий как бы подпирается листовым придатком, который в отличие от остальных, стерильных придатков является вильчатым. На первый взгляд может показаться, что синангии сидят в пазухе этих вильчатых придатков. В действительности каждый синангий расположен на верхушке короткой веточки, несущей вильчатый придаток. Эта веточка обычно короткая, и синангий кажется пазушным, но нередко она более или менее удлинена, а иногда (уже в виде аномалии) даже бывает повторно вильчато (дихотомически) ветвистая, с двумя или даже тремя синангиями - каждый на своей ножке. Эти и другие факты (в том числе ход проводящих пучков) свидетельствуют о том, что расположение синангиев верхушечное (терминальное) и что первоначально они сидели на концах более длинных веточек, которые в процессе эволюции укоротились. Интересно, что среди культивируемых в Японии клонов псилота голого есть один, известный под названием "Бунриу-зан" (Bunryu-san), вполне голый, т. е. лишенный листовых придатков, а синангии (состоящие обычно из более чем трех спорангиев) у него расположены на верхушках дихотомически разветвленных веточек. Если бы "Бунриу-зан" был обнаружен в девонских отложениях, то палеоботаники без колебаний отнесли бы его к риниофитам и, вероятно, поставили его где-то рядом с яравией.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'