Листья
Листья у мхов сидячие, большей частью поперечно прикрепленные к стеблю, простые, цельные, по краю могут быть зубчатыми и только очень редко глубоко разделенными (рис. 43).
На стебле листья расположены всегда по спирали, образуя двурядное или многорядное листорасположение. Закладываются листья в восходящей последовательности. Рост их начинается с деления двусторонней верхушечной клетки, от которой отделяются сегменты с двух сторон. После прекращения верхушечного роста листья достигают конечного размера посредством вставочного роста, происходящего в их основании.
По расположению на стебле различают листья низовые, срединные (или собственно стеблевые) и покровные.
Покровные листья окружают гаметангии и обычно отличаются от стеблевых размерами, формой, часто анатомическим строением, а иногда - окраской и консистенцией. Листья, окружающие архегонии, носят название перихециальных, а листья, окружающие антеридии,- перигониальных.
Низовые листья - сильно редуцированные, чешуевидные. Они развиваются в нижней надземной или в подземной части стебля у таких мхов, снабженных ризомом, как климациум (Climacium, табл. 8). Стеблевые листья расположены выше низовых, преимущественно в средней и верхней частях стебля.
Листовая пластинка чаще однослойная, реже целиком или частично дву-, многослойная. Клетки пластинки обычно богаты хлоропластами и выполняют функцию фотосинтеза. Жилка, если она развита, обычно проходит посередине листа и состоит из толстостенных, вытянутых клеток. Кроме механической роли, она обеспечивает проведение пластических веществ и воды.
По форме листья очень разнообразны (рис. 43). Чрезвычайно различно строение краев листьев. Завернутость и отвернутость краев листа создает микроскопические полости, где и удерживается капиллярная вода. Завернутый край листа, свойственный мхам, обитающим в условиях периодической засухи, выполняет еще и защитную функцию, предохраняя нежные части листа от иссушения.
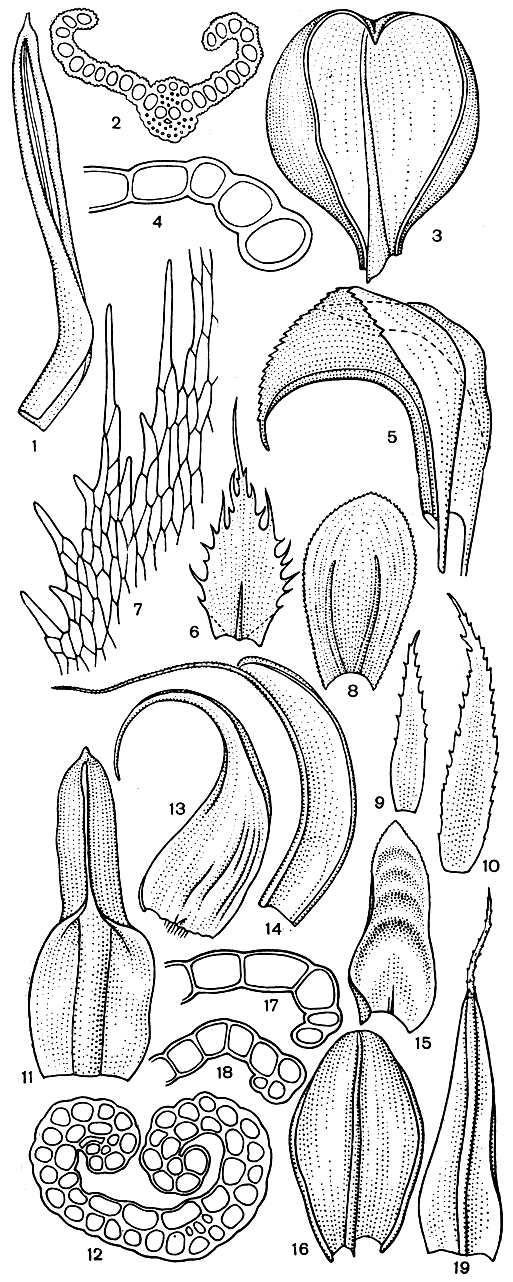
Рис. 43. Форма и поперечные срезы листьев бриевых. Вейсия (Weissia): 1 - лист с завернутыми краями; 2 - поперечный срез листа. Цинклидиум (Cinclidium): 3 - лист с сильно отвернутыми краями; 4 - срез через край листа. Палуделла (Paludella): 5 - оттопыренно отогнутый лист. Фаброния (Fabronia): 6 - лист с реснитчатым краем; 7 - часть края листа. Гукериопсис (Hookeriopsis incurva): 8 - широко яйцевидный лист с двойной жилкой. Эфемерум (Ephemerum): 9, 10 - листья без жилки. Индузиелла (Indusiella): 11 - лист с сильно завернутыми краями; 12 - поперечный срез листа. Гипнум (Hypnum): 13 - лист с двойной жилкой. Т ортула (Tortula): 14 - лист с бесцветным волоском. Неккерa (Neckera): 15 - языковидный поперечно-волнистый лист. Бриум (Bryum): 16 - яйцевидный лист; 17-18 - срезы через края листа. Гриммия (Grimmia): 19 - лист с волосковидным бесцветным кончиком
В листовой пластинке различают клетки двух основных типов: клетки паренхиматические - обычно округлые или угловатые до многоугольных (часто квадратные и шестиугольные), почти равной длины и ширины, и прозенхиматические - узкие, вытянутые в длину с заостренными, заходящими друг за друга концами.
У мхов вся пластинка листа редко бывает построена из одинаковых клеток. Если верхняя часть листа состоит из паренхиматических мелких клеток, более толстостенных, а иногда мамиллозных или папиллозных, то клетки листового основания почти всегда несколько крупнее и резко отличаются от верхних по форме.
Большой полиморфизм клеток мхов обусловлен не только разнообразием их формы, но и характером их клеточных оболочек. Здесь их многообразие, кажется, не знает пределов.
Кроме разрастания боковых стенок клеток пластинки листа, могут разрастаться и наружные стенки, образуя мамиллозность и папиллозность (рис. 43). Папиллы (утолщения клеточной оболочки) встречаются наиболее часто у бриевых мхов. Они бывают чрезвычайно разнообразной формы, а иногда к тому же в месте их образования клеточная оболочка разрастается и выпячивается. На этом образовании и сидят папиллы. Папиллы сильно увеличивают поверхность клеток, всасывающих воду, способствуют более быстрому собиранию воды и проведению ее внутрь клеток. Особенно часто развиваются папиллы на клетках у мхов сухих и сильно освещенных мест (тортула - Tortula, барбула - Barbula и др.), но широко представлены и у видов, обитающих в условиях избыточной влажности (у гелодиума шерстистого - Helodium lanatum, аулакомниума болотного - Aulacomnium palustre, у многих лесных мхов из семейства туидиевых - Thuidiaceae, у типичных эпифитов тропических и субтропических лесов из семейства метеориевых - Meteoriaceae).
У ряда мхов проявляется четкое обособление ассимиляционных и водоносных элементов листа. Это характерно для разных систематических групп мхов, далеко отстоящих друг от друга в родственном отношении. Наиболее ярко выраженный диморфизм клеток хорошо известен у сфагновых мхов, а из других листостебельных мхов - только у представителей семейства леукобриевых (Leucobryaceae) и калимперовых (Calymperaceae). Лист состоит из двух типов клеток: из зеленых, содержащих хлоропласты, и из мертвых, бесцветных, лишенных хлоропластов водоносных клеток.
У сфагновых мхов (рис. 44) хлорофиллоносные клетки развиты в большом количестве, окружают водоносные на протяжении всего листа. Водоносные клетки имеют спиральные и кольчатые утолщения и поры - отверстия в наружных оболочках. У леукобриевых в средней части многослойного листа расположено от одного до трех слоев хлорофиллоносных клеток (рис. 45). Почти у всех видов они со всех сторон окружены водоносными клетками с небольшим количеством пор преимущественно в основании листа.
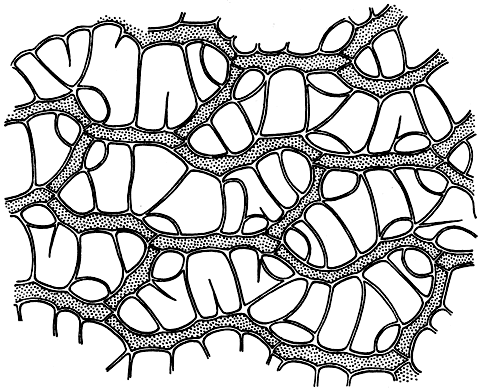
Рис. 44. Сфагнум болотный (Sphagnum palustre). Участок листа с клетками двух типов
По экологии и географии две названные группы мхов сильно отличаются. Сфагновые мхи - типичные обитатели болот, особенно широко распространенные в северном полушарии. К леукобриевым в основном относятся эпифиты тропиков и субтропиков, лишь некоторые виды леукобриума (Leucobryum) заходят в северную умеренную зону. Несмотря на это, у тех и у других выработались сходные приспособления в виде водоносных клеток.
Жилка у сфагновых мхов всегда отсутствует. У других верхоплодных мхов большей частью развита простая, т. е. неразветвленная, жилка. У бокоплодных мхов жилка даже в пределах одного семейства может иметь различный характер или отсутствовать. В этом отношении различают наиболее часто встречающуюся простую жилку; двойную, расходящуюся двумя лучами сразу же от основания листа; вильчатую, когда простая, не длинная жилка дает боковые, более короткие ответвления (рис. 43).
В большинстве случаев жилка выступает лишь на нижней, спинной стороне листа (рис. 43, 45) но у некоторых мхов также и на верхней. На спинной стороне жилки часто бывают разнообразные структурные образования (рис. 45). У видов рода фиссиденс (Fissidens) жилка на спинной стороне листа имеет крыловидный вырост. На верхней, брюшной стороне жилки у большинства политриховых находятся продольные, ассимиляционные пластиночки, состоящие из нескольких рядов клеток, богатых хлоропластами (рис. 46). Такие же пластиночки имеются у видов семейства даусониевых (Dawsoniaceae).
В семействе поттиевых у родов алоина (Aloina, рис. 45) и кроссидиум (Crossidium) в верхней части листа на жилке развиваются богатые хлоропластами многоклеточные, иногда разветвленные нити, которые часто подушковидно сплетаются.
Ассимиляционные пластиночки и нити в разных группах выполняют, по-видимому, одни и те же функции: у одних мхов - преимущественно ассимиляционную, у других - обеспечивающую главным образом всасывание воды. При подсыхании у большинства этих мхов края листьев заворачиваются и сверху плотно прикрывают пластиночки и нити. Обламывающиеся нити служат также для вегетативного размножения.
Большинство верхоплодных мхов имеет жилку сложного строения, состоящую из указателей, сопроводителей, стереидных пучков и наружных клеток (рис. 43, 45, 46).
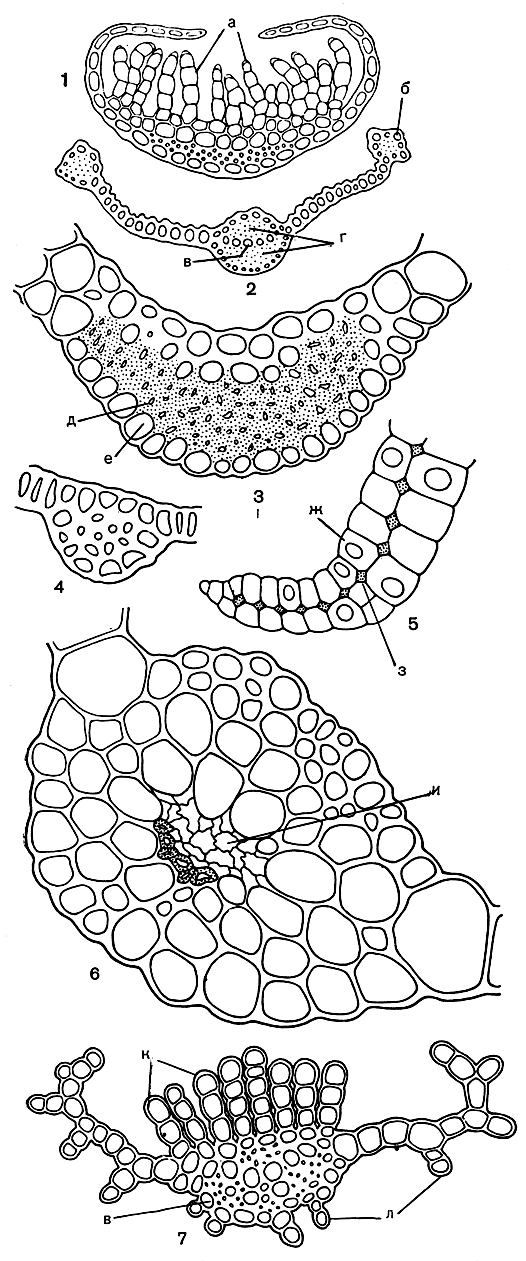
Рис. 45. Поперечные срезы листьев андреевых и бриевых: 1 - алоина (Aloina); 2 - сирроподон (Syrrhopodon); 3 - меезия трехрядная (Meesia trifaria); 4 - андрея Рота (Andreaea rothii); 5 - леукобриум (Leucobryum); 6 - мниум близкий (Milium affine); 7 - олиготрихум крылатый (Oligotrichum aligerum); а - ассимиляционные нити; б - край листа; в - указатели; г - брюшные и спинные стереиды; д - спинные стереиды; е - спинные клетки; ж - водоносные клетки; з - хлорофиллоносные клетки; и - сопроводитель; к - ассимиляционные пластиночки; л - спинныс пластиночки
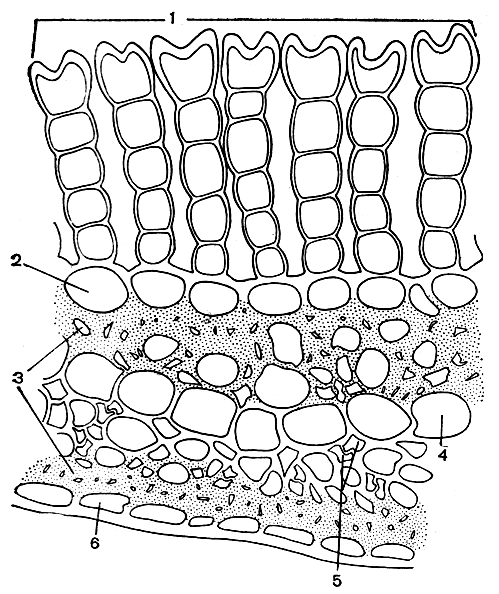
Рис. 46. Часть поперечного разреза пластинки листа политрихума обыкновенного (Polytrichum commune): 1 - ассимиляционные пластиночки; 2 - брюшные клетки; 3 - стереиды; 4 - указатели; 5 - гидроиды; 6 - спинные клетки
Паренхиматические клетки жилки листа с широким просветом, тонкостенные, бедные плазматическим содержанием, расположенные в один, реже в два ряда, называют указателями. Они проводят воду и на продольных стенках нередко снабжены порами. Состояние листа в основном зависит от тургора указателей. В сырую погоду они заполнены водой. Вследствие этого жилка, впитывая влагу, разбухает и вытягивается, а вместе с ней растягивается пластинка листа. В сухую погоду происходит обратное - жилка подсыхает, теряет свое натяжение, сгибается внутрь, а с нею изгибается и пластинка листа.
Сопроводители - тонкостенные, малого диаметра, вытянутые в длину клетки, объединенные в тяж и внешне сходные с клетками простого центрального пучка. Они почти всегда находятся на спинной стороне указателей.
Наружные клетки (обычно с довольно широким просветом) образуют как бы эпидерму жилки.
Стереиды жилки - толстостенные, вытянутые в длину и соединенные в тяжи клетки - сходны с лубяными волокнами сосудистых растений и по своему строению не отличаются от стереидных клеток стебля. Механическая ткань жилки образует один лентовидный стереидный пучок в середине жилки (рис. 45) или у ряда видов - два, спинной и брюшной, разделенные указателями (рис. 43, 45). Только у политриховых и даусониевых жилка имеет большое число близко расположенных стереидных пучков.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'