Стебель гаметофита
Все листостебельные мхи имеют более или менее развитое стеблевидное образование - каулидий, или стебель, покрытый листовидными выростами - листьями. У мхов установлены два основных типа роста стебля. Различают мхи ортотропные (от греч. orthos - прямой и tropos - поворот) с вертикальным ростом и плагиотропные (от греч. plagios - косой) с горизонтальным ростом стебля.
Характер роста стебля хорошо согласуется с расположением на стебле гаметангиев, в соответствии с чем выделяют верхоплодные (акрокарпные) мхи (табл. 6, 7) и бокоплодные (плеврокарпные) мхи (табл. 8). К верхоплодным принадлежат мхи с прямостоячими стеблями, на верхушке которых образуются архегонии, а со временем и спорогон. Вторая группа объединяет мхи со стеблями, плоско прижатыми к субстрату, свисающими у эпифитных мхов (табл. 8, 6), плавающими, как у фонтиналиса (табл. 8). У бокоплодных мхов архегонии закладываются на верхушке укороченных боковых побегов, и поэтому спорогоны сидят на боковых ветвях. Деление листостебельных мхов на верхоплодные и бокоплодные до некоторой степени является искусственным ввиду наличия переходных форм, но отражающим определенные тенденции в формативных процессах.

Таблица 6. Сфагновые мхи: 1 - сфагнум магелланский (Sphagnum magellanicum); 2 - сфагнум бурый (S. fuscum); 3 - сфагнум ленский (S. lenense); 4 - сфагнум Онгстрёма (S. aongstroemii); 5 - сфагнум балтийский (S. balticum); 6 - сфагнум дубравный (S. nemoreum); 7 - сфагнум оттопыренный (S. squarrosum); 8 - сфагнум береговой (S. riparium); 9 - сфагнум прорезной (S. perfoliatum)
В очертании на поперечном разрезе стебель чаще всего округлый, иногда овальный, угловатый или ребристый; последний образуется за счет сбегания жилки и краев основания листьев, которые, спускаясь по стеблю, срастаются с ним (рис. 42).
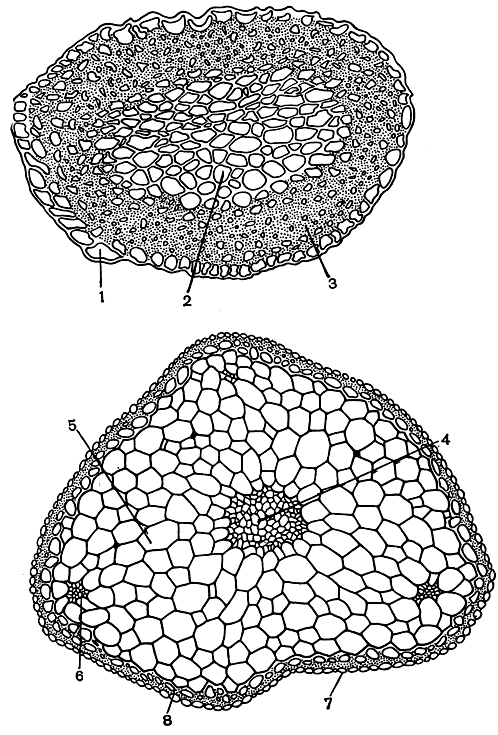
Рис. 42. Стебли бриевых на поперечном срезе. Вверху - пилотрихидиум (Pilotrichidium antillarum): 1 - гиалодерма; 2 - основная ткань; 3 - кора. Внизу - мниум (Mnium cinclidioides): 4 - центральный пучок; 5 - основная ткань; 6 - листовой след; 7 - гиалодерма; 8 - кора
Анатомическое строение стебля разнообразно. Стебель может быть построен из одинаковых клеток с толстыми или тонкими стенками или же из разных клеток. Во втором случае проявляется дифференциация на ткани: механическую, клетки которой по форме, утолщению стенок и их скульптуре сходны со склереидами сосудистых растений, и проводящие, обеспечивающие проведение воды и растворенных в ней минеральных веществ.
Механическую ткань, которая составляет кору, или склеродерму, стебля мхов, образуют стереиды - механические клетки стебля. Это вытянутые в длину, узкие, проземхимные клетки, обычно окрашенные в желтоватый, бурый, красно-бурый, пурпурный или почти черный цвет. Как правило, у этих клеток толстые стенки, причем оболочка у них иногда настолько утолщена, что просвет почти полностью исчезает. Во внутренних утолщенных частях стенок имеются водопроводящие поры. На внутренней поверхности стенок стереидных клеток могут образовываться утолщения в виде папиллообразных выростов, образующих густую сеть в полости клетки. Они служат для сохранения воды (клеточная оболочка способна к набуханию).
У ряда мхов один или несколько наружных слоев коры состоят из клеток, лишенных хлоропластов, с тонкими, прозрачными стенками и с более широким просветом (рис. 42). Эти слои клеток получили название гиалодермы (наружной коры). Гиалодерма развита у таких родов, как сфагнум, меезия (Meesia) и др. При подсыхании растения наружные стенки клеток гиалодермы впячиваются внутрь, а при поступлении воды стенки расправляются, клетки набирают воду и передают ее клеткам тканей, лежащих под гиалодермой.
С внутренней стороны кора граничит с основной тканью, которая состоит из однородных паренхимных клеток, часто заполняющих всю внутреннюю часть стебля (рис. 42).
Обычно кора постепенно переходит в основную ткань, реже кора резко отграничена от нее. Клетки основной ткани обильно заполнены плазматическим содержимым, хлоропластами, крахмалом, жирными маслами. Благодаря наличию хлоропластов основная ткань способна фотосинтезировать, но, кроме того, она служит местом сохранения воды, запасных веществ (крахмал, масла) и продуктов секреции (оксалат кальция). Основная ткань служит также для проведения воды.
У некоторых родов мхов в основной ткани обнаруживаются особые группы клеток, представляющие продолжение в стебле жилок листьев, по существу листовые следы. Различают два типа листовых следов: одни - слепо заканчивающиеся в основной ткани стебля и не доходящие до центрального пучка (обычно они состоят из группы одинаковых тонкостенных клеток), а другие - соединяющиеся с центральным пучком. Последние хорошо развиты в семействе политриховых (Polytrichaceae) в родах даусония (Dawsonia), сплахнум (Splachnum) и др. Листовые следы у политриховых имеют сложное строение, они дифференцированы на ткани, подобно стеблю этих мхов.
Наличие или отсутствие центрального пучка в большинстве случаев является критерием для разграничения видов, а иногда и родов. Центральный пучок (рис. 42), проходящий в середине стебля, состоит из вытянутых в длину, большей частью тонкостенных клеток с узким просветом, обеспечивающих проведение воды или ее сохранение.
Наиболее сложно построен центральный пучок у рода политрихум (Polytrichum) и у других представителей семейства политриховых, а также у рода даусония. В стебле политриховых можно наблюдать ткани из клеток гидроидов и лептоидов, напоминающие ткани стебля примитивных сосудистых растений (особое сходство обнаруживается со стеблем риниевых).
Гидроиды по характеру своей дифференциации, форме и отсутствию у них в зрелом состоянии цитоплазмы повторяют основные черты дифференциации и строения трахеид. По существу, единственное различие между теми и другими заключается в отсутствие у гидроидов лигнифицированных утолщений.
Лептоиды, расположенные на периферии тяжа гидроидов, во многом сходны с ситовидными элементами папоротниковидных. Сходство это проявляется в удлиненной форме и закругленных концах, а также в наличии в концевых и боковых стенках многочисленных отверстий.
В эпидерме стебля у всех мхов отсутствуют устьица. Кроме того, обычно из любой поверхностной клетки стебля могут возникать волосковидные образования - ризоиды - однорядные многоклеточные нити с бурыми оболочками и с косыми поперечными стенками. Обычно они служат для прикрепления к субстрату и адсорбции воды. Развиваются ризоиды главным образом при основании прямостоячего стебля, как, например, у фунарии (Funaria), или по всей длине ползучего или лежачего стебля на брюшной стороне, обращенной к субстрату. Ризоиды иногда покрывают весь стебель густым, светлым или окрашенным в бурый, фиолетовый или красноватый цвет войлоком; у некоторых мхов они скручиваются в длинные тяжи.
Ризоидный войлок обеспечивает поднятие из почвы воды в плотных дерновинках и более длительное ее сохранение в капиллярных пространствах между ризоидами и отдельными растениями. У водных мхов ризоиды выполняют главным образом функцию прикрепления к субстрату. На концах ризоидов у этих мхов образуются вильчатые разветвления, иногда сплетающиеся в подушечки, которыми растения прикрепляются к субстрату. При изменении условий питания и освещения ризоиды могут преобразовываться в зеленую вторичную протонему.
На верхушке стебля, в пазухе листьев, в местах отхождения листьев от стебля очень рано возникают своеобразные нитевидные образования из нескольких клеток, которые называют булавовидными волосками. Их булавовидная конечная клетка способна выделять слизь для защиты точки роста стебля от высыхания.
Кроме ризоидов и булавовидных волосков, на поверхности стебля у некоторых бокоплодных мхов развиваются пальчаторассеченные зеленые выросты стебля - парафиллии (от греч. para - возле и phyllon - лист). Иногда эти выросты образуются в таком количестве, что одевают стебель густым зеленым войлоком. Форма парафиллий чрезвычайно разнообразна. Они никогда не имеют жилок и расположены на стебле беспорядочно. Так же как и ризоиды, парафиллии способны не только проводить и удерживать воду, но и выполнять функцию фотосинтеза.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'