Эволюция проводящих клеток и тканей (А. А. Яценко-Хмелевский)
Возникновение покровной ткани, или эпидермы, было важным, но не единственным требованием при переходе растений к обитанию в наземных условиях. Как было показано выше, организация эпидермы с самого начала должна была отвечать двум прямо противоположным требованиям: с одной стороны, предохранять тело высшего растения от высыхания, с другой - позволять воздуху, содержащему углекислый газ, свободно достигать зеленых клеток с хлоропластами. Последнее, как мы помним, было возможным благодаря наличию устьиц. Свободно пропуская воздух, устьица также свободно пропускали воду, находящуюся в растении, что требовало ее постоянной подачи к зеленым частям растения. Так, на самых первых этапах развития высших растений выявилась одна из важнейших черт их физиологии - водный режим, т. е. совокупность всех тех процессов и структурных приспособлений, которые определяют поступление, передвижение и расходование воды.
Решение проблем водного режима привело к образованию ксилемы (от греч. xylon - дерево), или древесины,- системы тканей, специально приспособленных к выполнению функции водопроведения. Задача эта была не из легких. Дело в том, что проведение воды из одной живой клетки в другую осуществляется крайне медленно и требует определенных энергетических затрат. Рациональнее всего такая организация водопроводящей системы, при которой она состоит из мертвых клеток. Однако мертвая клетка может сохраняться среди живых тургоресцентных клеток только в том случае, если она будет достаточно механически укреплена. Такое механическое укрепление могло быть создано лишь при условии значительного утолщения клеточной оболочки и пропитывания ее веществом, придающим жесткость целлюлозному остову оболочки растительной клетки. Такое вещество в эволюции растений возникло очень давно, еще на заре жизни, до Появления высших растений. Это лигнин - сложное ароматическое соединение, чрезвычайно устойчивое к разрушающему действию Химических веществ и почти не поддающееся действию ферментов. В процессе эволюции растений лигнин первоначально возник, как полагают многие исследователи, в виде отброса в целях детоксикации (обезвреживания) некоторых ядовитых веществ, в частности фенольных соединений, образующихся в процессах жизнедеятельности клеток. Исключительно Удачное сочетание химических и физических свойств сделало лигнин одним из наиболее распространенных растительных веществ, стоящим на втором месте после целлюлозы у семенных растений: его содержание составляет около 60% от количества целлюлозы.
Оставим на некоторое время проблемы устройства самих элементов водопроводящей ткани и рассмотрим процесс их возникновения в теле высшего растения. По-видимому, у всех высших растений без исключения первые группы клеток водопроводящей ткани, а равно и флоэмы (от греч. phloios - кора, лыко) - ткани, проводящей растворы органических веществ, возникают из прокамбия. Прокамбий образуется из апикальной меристемы и является ее неотъемлемой частью, но все же это специализированная меристема, отличающаяся несколько вытянутой формой клеток и их цитологическими особенностями от остальных клеток апикальной меристемы.
Ксилему, образующуюся из прокамбия, называют первичной ксилемой. Она отчетливо подразделяется на два структурных типа - протоксилему, образующуюся на самых первых этапах дифференциации элементов ксилемы, и возникающую несколько позднее метаксилему. Отличия в строении этих двух типов мы опишем несколько позднее, а сейчас рассмотрим порядок образования элементов первичной ксилемы. Начнем с того, что прокамбий у различных растений возникает в верхушках осевых органов различными путями. Он может представлять собой на поперечном срезе небольшой кружок (а иногда и более сложную фигуру), лежащий в центре осевого органа, и, напротив, может развиться в виде небольшого пояска, к периферии и к центру от которого лежат неспециализированные клетки апикальной меристемы.
Формирование элементов первичной ксилемы из прокамбия в принципе может идти тремя различными путями. Оно может происходить центростремительно, т. е. самые первые элементы (протоксилема) возникают на периферий прокамбиального пучка, а самые последние (метаксилема) образуются в центре (или по крайней мере ближе к центру). Это экзархный тип образования ксилемы (или экзархная ксилема). Может быть и прямо противоположный тип формирования первичной ксилемы - когда вычленение элементов ксилемы из клеток прокамбия идет центробежно, от центра к периферии, когда, следовательно, протоксилема располагается в центре, а метаксилема образует внешний край первичной ксилемы. В таком - эндархном - типе ксилемы различают две его модификации. Если центробежное формирование ксилемы происходит в прокамбии, расположенном в виде колечка (и где, следовательно, центральная часть верхушки осевого органа превратится в сердцевину), то за таким типом образования первичной ксилемы сохраняют название эндархной ксилемы. Если же, напротив, центробежное формирование ксилемы происходит в прокамбии, представляющем собой один центральный пучок в верхушке осевого органа, то такую ксилему принято называть центрархной. Этот тип формирования ксилемы характерен для древнейших высших растений и справедливо считается самым примитивным. Напротив, эндархный тип рассматривается как наиболее совершенный.
Третий тип формирования ксилемы носит название мезархного. При этом самые первые элементы ксилемы возникают в средней части прокамбиального тяжа и последующее появление других элементов идет и к центру и к периферии возникающего тяжа первичной ксилемы.
Для подавляющего большинства семенных растений - всех покрытосеменных и большинства голосеменных - характерны два типа формирования первичной ксилемы, четко структурно отделяющих корень от стебля. В корне первичная ксилема формируется экзархно, а в стебле - эндархно. В низших отделах высших растений картина гораздо более разнообразна, и эти различия между экзархной, мезархной, центрархной и эндархной первичной ксилемой имеют часто очень большое систематическое и эволюционное значение.
Образованием первичной ксилемы может закончиться формирование водопроводящей ткани. При этом, как правило, весь прокамбий нацело дифференцируется в проводящие элементы ксилемы и флоэмы, и какие-либо новые клеточные деления, приводящие к образованию новых массивов проводящих тканей, оказываются невозможными. Такие растения обычно обозначают как "лишенные способности к вторичному росту", а их структуру называют первичной. В некоторых группах этого типа растений в массе тканей коры, уже закончивших свою дифференциацию и выполняющих функцию запасающей ткани, могут возникнуть новые очаги клеточных делений и возникающие вторичные меристемы дадут начало новым массивам проводящих тканей.
Однако у очень больших групп растений - всех хвойных и почти всех двудольных, а также некоторых бессеменных растений - в прокамбии между сформировавшейся флоэмой и ксилемой остается узкая полоска клеток, способных продолжать делиться. Так возникает камбий - латеральная (боковая) меристема, приводящая к образованию вторичных проводящих тканей - вторичной древесины и вторичной флоэмы. Именно они составляют огромные массивы тканей у всех древесных растений, а также большую часть ксилемы и флоэмы у травянистых двудольных.
А теперь вернемся к рассказу о том, как ведут себя элементы водопроводящей ткани в процессах своего превращения из прокамбиальной или камбиальной клетки в высокоспециализированную клетку ксилемы.
Процесс отложения вторичной целлюлозной оболочки и последующее пропитывание ее лигнином осуществляется еще в живой клетке ее живым протопластом. При этом на первых этапах развития молодого растения (или молодых частей старого растения) все клетки растущих частей растения удлиняются. Однако такое удлинение клеток практически невозможно при наличии сплошной жесткой, пропитанной лигнином оболочки. В этом отношении, опять-таки на самых первых этапах развития высших растений, было выработано оптимальное приспособление, сохранившееся практически у всех растений на всех этапах последующей эволюции,- появление кольчатых и спиральных водопроводящих элементов, т. е. клеточных элементов, у которых вторичная оболочка не одевает сплошь всю первичную оболочку, а расположена в ней кольцами или спиралью (рис. 6). Подобное решение позволяет молодому растению (или молодой части растения) иметь растущие в длину клетки, но с очень обедненным содержимым и хорошо сопротивляющиеся давлению соседних живых клеток. Процесс этот бывает хорошо заметен при рассматривании под микроскопом участков тела растения различного возраста. У очень молодых проростков кольца или витки спирали таких водопроводящих клеток сближены, тогда как у более зрелых они расходятся. Это показывает, что отдельные участки первичной оболочки между лигнифицированными (одревесневшими) кольцами или витками спирали продолжают свой рост.
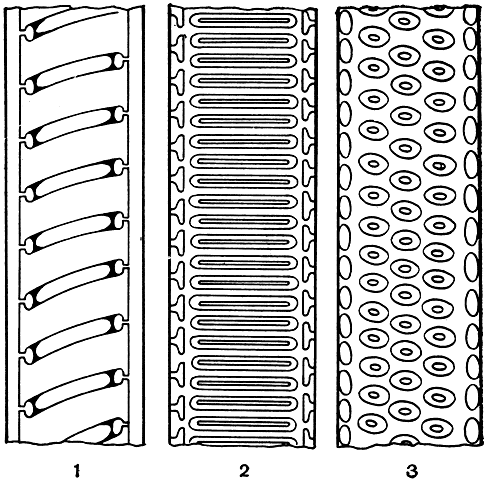
Рис. 6. Основные типы трахеид: 1 - спиральные; 2 - лестничные; 3 - точечные
Спиральные или кольчатые элементы могут быть цельными клетками, полностью одетыми первичной оболочкой, или же иметь на своих концах сквозные отверстия - перфорации. В первом случае такие элементы обозначают как кольчатые или спиральные трахеиды, во втором - как кольчатые или спиральные членики сосуда. К определению этих двух названий мы вернемся несколько позже, а пока еще раз подчеркнем, что такие спиральные и кольчатые элементы являются самыми первыми как в эволюции высших растений, так и в индивидуальном развитии каждого отдельного растения (в его онтогенезе). Это не случайно. Действительно, такой тип вторичного утолщения и самый экономный и вместе с тем чрезвычайно целесообразный в молодых, только что образовавшихся частях растения. У некоторых растений - многих риниофитов, а также у более подвинутых по пути эволюции таксонов в случае редукции их тела - этот тип водопроводящих элементов является не только первым, но и единственным. Несколько забегая вперед, скажем, что кольчатые и спиральные элементы в огромном большинстве случаев образуются из прокамбия, причем на самых первых порах его дифференциации (т. е. путем превращения этих образовательных тканей в постоянную ткань).
При всех достоинствах кольчатых и спиральных элементов как путей для проведения воды их механическая прочность оставляет желать лучшего. Поэтому, после того как у молодого растения (или молодой части растения) закончились ростовые процессы, связанные с удлинением клеток, в ксилеме начинают формироваться элементы со сплошной вторичной одревесневшей оболочкой. Такие клетки, как только формирование клеточной оболочки в них бывает закончено, совершенно не способны удлиняться (или расширяться) и очень быстро отмирают. Процесс отмирания происходит в них в весьма организованной форме.
В живом протопласте среди прочих органелл имеются специальные тельца - лизосомы, внутри которых находятся ферменты, расщепляющие вещества, слагающие протопласт. Как при этом они не растворяют оболочку самой лизосомы, до сих пор не ясно. Когда клетка завершила образование вторичной оболочки и протопласт становится уже помехой в выполнении ее основной функции - проведении воды, лизосома вскрывается, заключенные в ней ферменты выходят наружу в цитоплазму и начинается стремительный процесс растворения протопласта. Остатки его вымываются током воды, и в очень короткий срок клетка оказывается не только мертвой, но и пустой.
Вторичная оболочка вокруг водопроводящего элемента, разумеется, не может быть совершенно сплошной, в ней всегда бывают углубления, доходящие до первичной оболочки. Эти углубления получили в русской ботанической литературе чрезвычайно неудачное название - поры. Под "порой" и в технической литературе, и в обыденной жизни понимают сквозное отверстие, тогда как пора в анатомическом смысле - это не дырка, не отверстие, а только углубление в толще вторичной оболочки, или ямка. Неудовлетворительность этого названия осознавали многие русские анатомы растений, но сила привычки оказалась сильнее, и термин этот все-таки закрепился. В английской ботанической литературе употребляется более удачный термин pit, что в переводе означает яма, углубление, впадина.
В огромном большинстве случаев водопроводящие элементы несут своеобразные поры - окаймленные (рис. 7). Принцип устройства окаймленной поры заключается в том, что канал, открывающийся в полость клетки, более или менее внезапно расширяется, образуя то, что называют камерой поры. Дном ее является первичная оболочка. Такая пора в одной проводящей клетке, как правило, точно совпадает с такой же порой у соседней клетки, так что образуется пара пор, соединяющая две клетки.
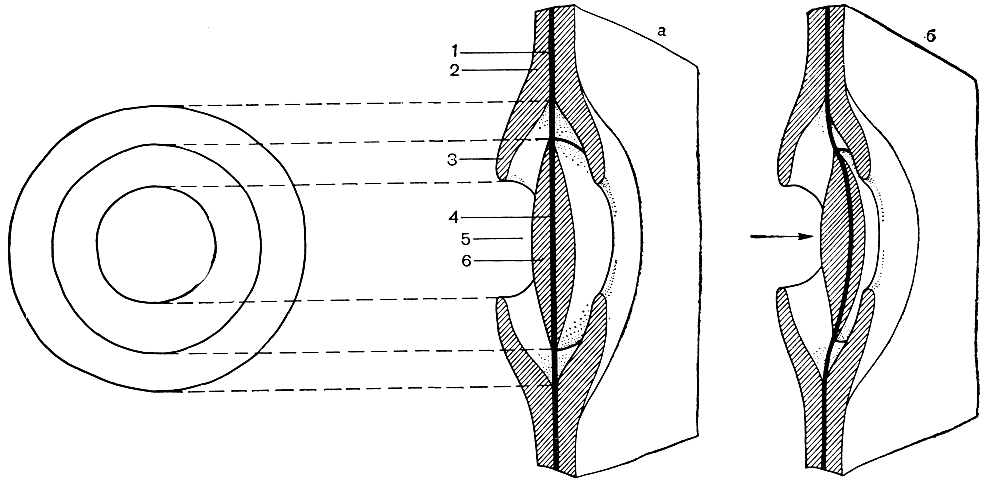
Рис. 7. Схема строения пары окаймленных пор: 1 - первичные оболочки двух соседних клеток (и межклеточный слой между ними); 2 - вторичная оболочка; 3 - поровое окаймление; 4 - поровая мембрана (состоящая из двух первичных оболочек соседних клеток и межклеточного слоя между ними); 5 - поровая камера; 6 - торус: а - открытое положение поровой мембраны, б - закрытое положение
Окаймленные поры - очень характерная особенность водопроводящих клеток практически у всех высших растений. Они могут иметь различное строение, могут и располагаться по-разному на стенках водопроводящих клеток. Все эти детали имеют большое значение и в систематике высших растений, и для понимания их эволюции.
Водопроводящие клетки, имеющие замкнутую со всех сторон первичную оболочку, на которой наслоена вторичная оболочка, располагающаяся в виде спиралей или колец либо пронизанная окаймленными порами, называют трахеидами (от греч. tracheios - горло и eidos - вид). Трахеида - это основной водопроводящий элемент высших растений с момента их появления на суше и по сей день. Вместе с тем замкнутая клетка, из которой вода - должна переходить в соседние клетки через окаймленные поры, просачиваясь через две первичные оболочки (и через склеивающий их межклеточный слой), не является оптимальным решением для быстрой подачи воды. Действительно, в процессе эволюции в ряде групп высших растений возник новый проводящий элемент - членик сосуда. Для члеников (или элементов) сосуда характерно наличие в концах клеток перфорации. Соединяясь между собой такими перфорациями, сотни, тысячи и десятки тысяч члеников образуют сосуды - длинные сквозные трубки, которые, соединяясь между собой стенками с окаймленными порами, тянутся вдоль оси растения, по корню и стеблю, проводя воду от кончиков корней до верхушек ветвей и жилок листьев.
В строгой анатомической терминологии сосудом называют вертикальный ряд члеников сосудов, а члеником сосуда называют только клетку, несущую две (или иногда одну) перфорации. Иногда сосуды или членики сосудов называют трахеями. Это, по существу, устаревшее название было предложено еще в XVII в. выдающимся итальянским естествоиспытателем Марчелло Мальпиги, впервые описавшим сосуды в древесине груши и принявшим их за дыхальца, аналогичные трахеям насекомых. Последующие авторы, наблюдавшие трахеиды в древесине хвойных, поняли отличие их от члеников сосудов, но, установив их аналогичную функцию, дали им название "сосудоподобных", т. е. трахейд. Последний термин укрепился и в настоящее время является строго стандартным.
По внешнему виду можно различать два типа пор на стенках трахеид - более или менее правильно округлые и сильно вытянутые (перпендикулярно длинной оси клетки), в последнем случае совокупность пор на стенке трахеиды напоминает лестницу, что обозначается термином лестничная поровость. Названное различие между округлыми и вытянутыми порами, как это ни странно, является весьма существенным. В обширной группе кордаитовых и хвойных, которые известны с начала карбона и до наших дней и которые характеризуются сильным развитием древесины, составленной почти исключительно из трахеид, никогда не встречаются трахеиды с лестничным расположением пор. Напротив, у папоротников и таких голосеменных, как семенные папоротники, беннеттитовые, саговниковые и некоторые другие,
Для большинства хвойных характерно особое устройство окаймленной поры: наличие так называемого торуса, т. е. утолщенного участка первичных оболочек в центре дна поры - на пленке поры (рис. 7). Торус способен работать как клапан, закрывая отверстие канала поры и тем самым выключая трахеиду из водного тока. Однако этот клапан только одноразового действия: закрыв пору, он больше не способен прийти в центральное положение и вновь включить трахеиду в водный ток. Тем не менее это весьма существенное физиологическое приспособление, поскольку способность выключать часть водных путей после утраты в них надобности очень важна для растения.
Водопроводящие элементы, обладающие достаточно толстой оболочкой, инкрустированной лигнином, играют в теле растения также и механическую роль. Это совмещение роли водопроведения с ролью механической особенно существенно в классе пинопсид (кордаитовые и хвойные). Однако механические производные водопроводящих элементов, о которых будет речь ниже, четко выражены лишь у цветковых растений. Для этих последних характерна особенно высокоспециализированная водопроводящая система, основным элементом которой являются членики сосудов.
Нужно учитывать, что появление члеников сосудов, т. е. продырявленных водопроводящих элементов, имело место в эволюции высших растений несколько раз, и сосуды были отмечены у ряда видов селагинелл, хвощей, некоторых папоротников, а также голосеменных (класс гнетопсиды). В соответствующих разделах настоящего тома "Жизни растений" о всех этих случаях будет сказано подробнее. Но только у цветковых растений сосуды стали неотъемлемой частью ксилемы и, по-видимому, сыграли большую роль в эволюции этой самой подвинутой и самой процветающей группы высших растений. Примечательно, что первые цветковые растения были еще бессосудисты и в их ксилеме имелись только трахеиды, в основном лестничного типа. Такие гомоксильные (т. е. с однородной ксилемой) покрытосеменные дожили до наших дней.
Членики сосудов у цветковых растений морфологически довольно разнообразны и образуют очень четкий эволюционный ряд, начинающийся от члеников с многочисленными лестничными перфорациями и до члеников с простыми перфорациями, расположенными на поперечных стенках.
На рисунке 8 изображен широко известный эволюционный ряд члеников сосудов покрытосеменных. Основанный на работах выдающегося американского ботаника И. Бэйли и его школы, этот ряд всегда помещается во всех учебниках как одна из наиболее бесспорных последовательностей эволюционных преобразований такого важного физиологического элемента, как водопроводящая клетка. Действительно, даже без экспериментальных подтверждений (а такие есть) и без гидродинамического расчета (который проводился) можно усмотреть усовершенствование этих отдельных элементов длинной трубки - сосуда. Совершенно очевидно, что вода с большей легкостью идет по трубке, где отверстия расположены на поперечных стенках члеников, чем по трубке, в которой отверстия расположены по бокам, или тем более по трубке, разделенной по всей длине мелкими перекладинами.
Следует только иметь в виду, что совершенство водопроводящего аппарата у высших растений далеко не всегда является решающим фактором в борьбе за существование. В условиях высокой влажности и в затененных местах способность быстро проводить воду часто не имеет особого или вовсе никакого значения. Напротив, в местах с низкой относительной влажностью воздуха, где высокая температура и редкие осадки создают для растения напряженную ситуацию, совершенство основного элемента ксилемы - сосудов - может играть решающую роль в выживании. Поэтому, как относительно недавно установлено, существует четкая зависимость между сухостью местообитания и процентом растений с лестничными перфорациями: чем выше сухость, тем процент этот ниже, а в засушливых (аридных) условиях он, как правило, равен нулю.
Столь же важна эффективная водопроводящая система и у тех древесных растений, которые высоко выносят свою листву, поднимая ее ближе к солнечному свету. В соответствии с этим у огромного большинства древесных растений первого яруса, даже в тропическом дождевом лесу, обязательно встречаются сосуды с простыми перфорациями, в то время как у видов нижних ярусов преобладают или встречаются у достаточно большого количества видов лестничные перфорации. То же самое относится и к лианам. Вынужденные тянуть воду часто на протяжении десятков метров, лианы всегда характеризуются сосудами с простыми перфорациями.
Сказанное относится не только к цветковым растениям. Как было установлено еще в прошлом столетии, у папоротника орляка тоже встречаются настоящие сосуды. По внешнему виду перфорации сосудов орляка очень напоминают лестничную поровость трахеид других папоротников, но у орляка дно (пленка) пор исчезло и возникло сквозное отверстие. Многие ученые связывают широкое распространение орляка по всему северному полушарию и способность его заселять относительно засушливые местообитания именно с наличием у него сосудов, а не трахеид. Правда, сосуды имеются и у некоторых других папоротников.
Мы уже говорили, что сочетание в одном клеточном элементе - трахеиде - и способности проводить воду, и механической прочности было важным эволюционным приспособлением. Появление сосудов с их все увеличивающейся полостью и относительно тонкими стенками несколько ослабило бы механическую роль ксилемы, если сосуды остались бы единственным элементом этой ткани. Решение этой проблемы у разных растений было неодинаково. Для настоящих древесных растений, у которых основная масса ствола и корня составлена древесиной, решение могло быть только одно: в древесине (ксилеме) было необходимо сохранить и другие, кроме сосудов, элементы, у которых водопроводящая функция была бы ослаблена или даже сведена к нулю, тогда как возрастала бы функция механическая. Рисунок 9 показывает такой же эволюционный ряд, что и на предыдущем рисунке 8, но идущий уже в другом направлении - от все той же трахеиды, которая начинает собой эволюционный ряд члеников сосудов, к строго механическому элементу - волокну либриформа(от лат. liber - луб) - через промежуточную стадию волокнистой трахеиды.
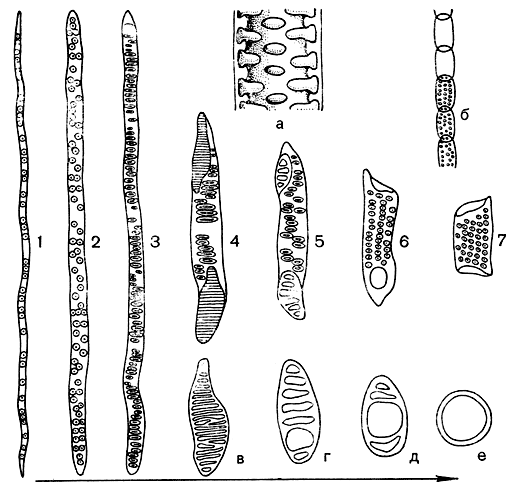
Рис. 8. Изменения структуры трахеальных элементов в ходе эволюции: 1, 2 - трахеиды с округлыми окаймленными порами; 3 - трахеиды с вытянутыми окаймленными порами; 4-7 - членики сосудов, характеризующиеся различной степенью изменений в направлении сокращения их длины, уменьшения угла наклона конечных стенок, преобразования лестничных перфораций в простые; а - фрагмент продольного среза трахеального элемента, б - расположение члеников в сосуде, в-е - типы перфораций между члениками сосудов лестничные трахеиды вполне обычны, хотя, впрочем, также обычны и трахеиды с округлыми порами

Рис. 9. Изменения структуры ксилемных волокон в ходе эволюции. Изменения состоят в сокращении длины волокон, в уменьшении размеров окаймления и отверстий пор
Здесь совершенно отчетливо прослеживаются два направления: с одной стороны, утолщение оболочек и, с другой, - все большая редукция окаймления пор, которое полностью исчезает у либриформа. Поры у волокон либриформа видны в виде узкой щели, вокруг которой нельзя выявить никакого окаймления. Этот процесс редукции настолько постепенен, что, для того чтобы отличить волокнистые трахеиды от волокон либриформа, еще сорок лет назад было принято решение считать все поры волокон простыми, если при увеличении в 500 раз окаймление у них незаметно. Древесина, в которой наряду с сосудами имеется большее число волокон либриформа, естественно, приобретает высокую механическую прочность, что, в свою очередь, позволяет растению развивать очень высокий ствол, несущий обширную крону.
Подчеркивая значение механической функции древесины, следует сказать, что ее роль в этом отношении не всегда была столь решающей, как у современных деревьев. Первые растения суши - риниофиты - отличались незначительными размерами, но их ксилема, составленная в значительной степени из кольчатых и спиральных трахеид, образовывала тонкий цилиндр, расположенный в центре оси, и все же играла некоторую механическую роль. Относительно скоро эволюция привела к появлению самых различных групп высших растений, которые можно, однако, разделить на две условные группы по той роли, которую в них играет ксилема. К первой из этих групп относятся бессеменные высшие растения и некоторые семенные. Для всех древовидных их представителей характерно мощное развитие живых паренхимных тканей, составляющих всю основную массу ствола и корней (или заменяющих их органов). Древесина у этих растений образует, как правило, узкий центральный цилиндр или узкое кольцо, обрамляющее мощную сердцевину. Даже когда эти растения достигают значительных размеров, как некоторые современные саговниковые или древовидные папоротники, все же основная масса их тела представлена живыми паренхимными клетками. Напротив, кордаитовые и хвойные, а также древесные двудольные характеризуются мощным развитием древесины, объем которой превосходит объем всех остальных тканей осевых органов растения.
Различие это, по-видимому, играет в эволюции высших растений довольно существенную роль. Дело в том, что для создания мощной древесины (типа древесины хвойных или типичных древесных двудольных) требуется чрезвычайно высокая координация деятельности образовательных тканей растения, что, в свою очередь, возможно только при достаточно сложной системе взаимодействия физиологически активных веществ. Мы уже говорили о том, что у всех высших растений рост в длину (в высоту) достигается за счет постоянных клеточных делений в участках тела растения, расположенных на окончаниях стебля и корня и их разветвлений. Это апикальные меристемы (рис. 10). Они работают практически в течение всей жизни растения. Действительно, растение, в отличие от животных, всю свою жизнь растет, т. е. постоянно наращивает новые участки к своим надземной и подземной частям.
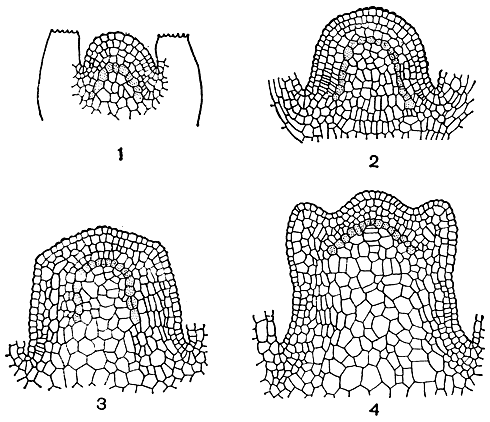
Рис. 10. Верхушечные меристемы побега на примере зверобоя уральского (Hypericum uralum): 1 - заложение пары листьев; 2 и 3 - постепенное увеличение и изменение формы верхушки побега; 4 - заложение следующей пары листьев
Русский язык словом растение хорошо подметил эту их особенность. Аналогичный термин (Gewächse) есть и в немецком языке, также связанный с понятием роста (Wachstum). Латинский, английский и французский языки (planta, plant, plante) подчеркнули в названии "растение" другую, тоже весьма характерную черту - прикрепленность, "посаженность" в почву. Однако постоянный рост - примечательнейшая черта любого высшего растения.
Апикальные меристемы не способны привести к значительному утолщению оси растения. Разумеется, клетки, после того как они выделились из меристематической зоны и начали принимать свою окончательную форму, увеличиваются в объеме и тем самым увеличивают и размеры тела растения. Увеличение это, однако, никогда не может быть очень значительным, и, самое главное, оно однократно: раз достигнув определенного диаметра, ось растения перестает расти. Такое ограничение в росте по диаметру в большинстве случаев для растения невыгодно, и в процессе эволюции у самых различных групп высших растений стали появляться очаги клеточных делений, расположенные не в конечных разветвлениях оси, а вдоль нее, латерально (по бокам). Как мы уже знаем, это - камбий, или латеральная меристема. Деятельность камбия может быть достаточно неупорядоченной, он может работать некоторое время и потом навсегда замирать, и все слагающие его клетки при этом теряют способность к делениям. При этом очень часто в более внешних слоях возникают новые участки клеточных делений, и так несколько раз.
Клетки, возникающие в результате делений в камбии, могут образовывать различные ткани и чаще всего проводящую ткань. Развитие мощной древесины как раз и характеризуется высокой упорядоченностью клеточных делений в камбии, приводящих к образованию значительных масс ксилемы. При этом возникает типичное дерево, такое, каким являются привычные для нас сосна или ель, береза, тополь или дуб. Привычность понятия "дерево" как типа организации тела растения (этой его "жизненной формы") обычно приводит к мысли о его простоте. Однако оказывается, что дерево для своего существования требует очень сложных связей между отдельными группами делящихся клеток, обеспечивающих ту регулярность работы камбия и апикальных меристем, которые и создают это стройное чудо природы - дерево.
Если утеря той гармоничной согласованности, которая характерна для дерева, происходит довольно легко, достоверные случаи ее восстановления нам вообще неизвестны. Разумеется, многие травы в процессе эволюции дают вновь древовидные формы. Это отмечается и среди маревых (вспомним саксаул), и у сложноцветных, и во многих других семействах. Но, однако, при этом не удается вновь найти ту регулярную форму организации, которая характерна для настоящих деревьев и которая способна у одной и той же особи работать "как часы" в течение долгих десятилетий и столетий. Надо помнить также, что во всех тех районах земного шара, где недостаток влаги или тепла не ограничивает рост растений, произрастают леса, в которых по биомассе и физиономически всегда господствуют именно настоящие деревья, простота строения которых достигается высочайшей (для растения) физиолого-биохимической организацией.
В очерке об основных признаках строения высшего растения мы значительное внимание уделили водопроводящим тканям. Это не случайно. Помимо того, что водный режим является одной из важнейших физиологических функций высшего растения, без нормального осуществления которой невозможно выполнение всех других его физиологических отправлений, одревесневшие клетки, слагающие ксилему, отличаются исключительной стойкостью к самым различным неблагоприятным воздействиям. Поэтому древесина превосходно сохраняется в ископаемом состоянии, давая палеоботаникам отличнейший материал для исследования. Она сохраняется также в теле многолетних растений в течение всей их жизни, часто продолжающейся десятилетия или столетия, а у некоторых растений - даже тысячелетия. Именно эти обстоятельства и определяют интерес к древесине со стороны систематиков, палеоботаников и экологов, изучающих жизнь растений с самых различных сторон.
Заканчивая раздел о водопроводящих тканях высших растений, надо отметить, что у тех растений, где древесина образует значительные массивы клеток, в ее составе, помимо чисто водопроводящих и механических элементов, всегда имеется некоторое количество и живых клеток. В огромном большинстве случаев эти живые клетки характеризуются также лигнифицированными (одревесневшими) клеточными оболочками и по своим механическим свойствам мало отличаются от водопроводящих и механических элементов древесины (эти два последних и важнейших типа клеток древесины часто объединяют общим термином трахеальные элементы). Живые элементы древесины в большинстве своем не вытянуты вдоль оси ствола (или корня) и имеют более или менее изодиаметрическую (равностороннюю) форму. Поэтому их обычно называют паренхимой древесины, поскольку в анатомии растений под термином "паренхима" понимают ткани, состоящие именно из изодиаметрических клеток.
В массе мертвых водопроводящих клеток сохраняется какое-то (иногда довольно значительное - до 25% и более от общего объема древесины) количество живых элементов, что, по-видимому, объясняется необходимостью где-то откладывать запасные вещества, так как у деревьев нет специальных запасающих органов, столь характерных для многих травянистых растений. Существуют и другие объяснения наличия живых тканей среди массы мертвых клеток. Как бы то ни было, паренхима в древесине существует, что еще более усложняет строение этой ткани.
Паренхима в древесине существует, как правило, в виде двух модификаций. С одной стороны, у очень многих древесных растений имеются клетки паренхимы, собранные в вертикальные тяжи, тянущиеся вдоль оси дерева, - это тяжевая, или древесинная, паренхима. Вместе с этой паренхимой у огромного большинства древесных растений имеются группы живых клеток, собранных в горизонтальные полосы. Такие полосы получили название сердцевинных или ксилемных лучей. Вся система живых клеток образует сложную сеть, причем отдельные звенья этой сети обычно на том или ином уровне соприкасаются друг с другом. Наиболее сложно паренхимная система устроена у цветковых растений, и мы будем иметь случай вернуться к ней в соответствующих разделах настоящего издания.
Проводящие ткани растения не исчерпываются лишь водопроводящей тканью - ксилемой, по которой проходит ток воды от корней к листьям. Наряду с ней существует ткань, проводящая органические вещества, образованные в результате процесса фотосинтеза в зеленых клетках листа, от кроны вниз, к корням (а также, конечно, к цветкам и плодам). Это - флоэма, или луб, по которому идет нисходящий ток, противопоставляемый току воды с растворенными минеральными солями - восходящему току.
В отличие от некоторой простоты функционирования водопроводящих элементов, работа которых с известной точностью может быть описана в терминах гидродинамики и в целом давно уже не представляет собой физиологической загадки, движение веществ по специализированным клеткам флоэмы (ситовидным трубкам) до сих пор еще не получило объяснения, приемлемого для всех специалистов в области "флоэмного транспорта". Сложность проблемы заключается в том, что растворы Сахаров и аминокислот передвигаются по флоэме с относительно высокой скоростью (около 1 м в час). Эта скорость несравненно меньше, чем скорость движения воды по ксилеме (весьма различная у разных растений, но достигающая иногда нескольких десятков метров в час), но тем не менее в тысячу раз большая, чем если бы вещества по ситовидным трубкам "перетекали" путем диффузии. Скорость тока веществ по флоэме устанавливается в относительно несложных экспериментах с применением меченых атомов.
Так же как и элементы ксилемы, элементы флоэмы в процессе эволюции высших растений претерпели целый ряд значительных морфологических и функциональных изменений. Однако надо подчеркнуть, что если водопроводящие элементы явились некоторым новым приобретением у первых высших растений, осуществивших выход на сушу, то элементы, аналогичные флоэме, уже имелись у некоторых водорослей, и, таким образом, в эволюционном плане флоэма значительно старше ксилемы. Более того, можно сказать, что основной физиологический признак клеток флоэмы - их способность проводить растворенные органические вещества - в той или иной степени вообще присущ любой живой клетке, и, следовательно, механизмы, обеспечивающие эту способность, возникли на самых первых этапах развития жизни на Земле. Само собой разумеется, при выделении отдельных групп клеток, специально приспособленных для проведения органических растворов, эта способность была развита и улучшена, но по сути своей оставалась той же. Она позволяла бактериальной клетке или клетке сине-зеленых водорослей передвигать растворы в пределах своего крошечного тельца. В этом отличие клеток флоэмы от клеток ксилемы, которые, чтобы дать им возможность выполнять свою функцию в жизни наземного растения, было необходимо прежде умертвить.
Рассматривая эволюцию флоэмы, нужно отметить, что тонкие целлюлозные оболочки ее клеток несравненно хуже сохраняются в фоссилизированном ископаемом состоянии, чем клетки ксилемы с ее одревесневшими стенками. Поэтому наши сведения о последовательных этапах развития элементов флоэмы, столь подробно изученных для ксилемы, гораздо более отрывочны. По имеющимся данным можно предположить, что у самых первых наземных растений - риниофитов - клетки флоэмы отличались от других паренхимных элементов (фотосинтезирующих или запасающих) главным образом своей вытянутой вдоль оси формой. Однако даже такие просто устроенные растения, как псилот и тмезиптерис, уже имеют на стенках ситовидных элементов ту структуру, которая и дала название этому типу клеток, а именно собрание мелких, своеобразно устроенных отверстий, образующих "ситечко" ("ситовидный участок", или "ситовидное поле" по более строгой терминологии). Впрочем, несмотря на свою несомненную глубокую древность, и псилот и особенно тмезиптерис представляют собой, как об этом будет подробнее сказано в главе о псилотовидных, достаточно специализированные растения, и вполне вероятно, что присущие им ситовидные элементы являются результатом длительной эволюции.
Высокоспециализированные поры, совокупность которых образует ситовидное поле (или - у покрытосеменных - ситовидную пластинку), представляют собой, по сути дела, расширенные плазмодесмы - тончайшие тяжи цитоплазмы, пронизывающие первичную клеточную оболочку и связывающие между собой протопласты соседних клеток. Плазмодесмы - чрезвычайно тонки и, за редчайшими исключениями, могут быть замечены только на экране электронного микроскопа. Как правило, дно простой поры (точнее, первичного норового поля, поскольку именно этим, несколько неуклюжим термином обозначают обычно поры в первичной оболочке) пронизано большим (до десяти) числом плазмодесм. Поры ситовидных полей заполнены одним цитоплазменным тяжем, размеры которого в десятки раз превышают размеры плазмодесмы соседних клеток. Насколько можно судить по имеющимся разрозненным данным, такая огромная плазмодесма возникает в результате слияния многих отдельных плазмодесм. Впрочем, нет уверенности, что это единственный путь образования пор ситовидных элементов и, возможно, что у некоторых растений гигантские плазмодесмы пор ситовидных элементов образуются просто путем расширения начальной тонкой плазмодесмы.
В классическом случае, канал поля ситовидного элемента, заполненный цитоплазмой, облицован особым углеводом - каллозой, которая покрывает цитоплазменный тяж чехлом. Кроме того, у многих растений каллоза окружает небольшим валиком отверстия канала.
Специализированные поры встречаются, по-видимому, у всех высших растений (за исключением, быть может, риниофитов), но тот порядок, в котором они собраны в ситовидные поля, различен у разных групп. По-видимому, у всех так называемых "споровых растений" (хвощевидных, плауновидных, папоротниковидных), так же как и у голосеменных, такие каналы собраны в отдельные, часто небольшие группы, рассеянные по всей длине оболочки ситовидного элемента. Напротив, у цветковых растений ситовидные поля собраны в более или менее крупные группы, обычно обозначаемые как ситовидные пластинки (рис. 11, 12). Ситовидные элементы, в которых каналы собраны в ситовидные пластинки, обозначаются уже как ситовидные трубки. В пределах этого типа ситовидных элементов различаются разные морфологические структуры, как мы об этом скажем чуть ниже. Эти структурные различия между ситовидными элементами всех остальных высших растений и ситовидными трубками покрытосеменных обычно аналогизируются с различиями между трахеидами и члениками сосудов. Для этой аналогии есть несомненные основания, но различия между члениками сосудов и трахеидами более четкие.
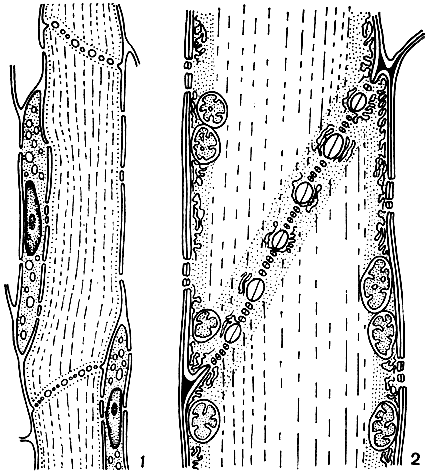
Рис. 11. Структура ситовидных трубок флоэмы: 1 - расположение ситовидных элементов и клеток-спутниц в ситовидной трубке; 2 - фрагмент продольного среза ситовидной трубки (район ситовидной пластинки)
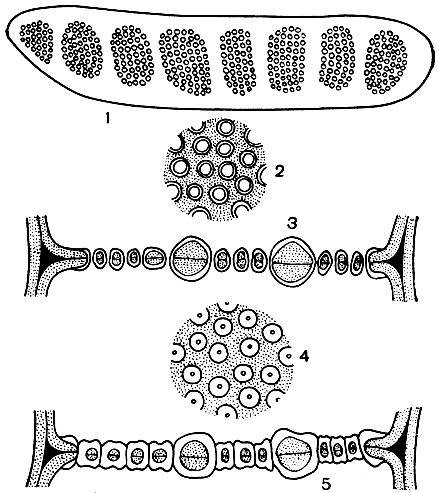
Рис. 12. Структура ситовидных трубок флоэмы: 1 - вид сверху сложной ситовидной пластинки; 2 и 3 - ситовидная пластинка на поперечном и продольном срезах ситовидной трубки при низком содержании каллозы в порах; 4 и 5 - то же самое при высоком содержании каллозы
Чрезвычайно важным признаком организации ситовидных элементов является отсутствие в этих клетках клеточного ядра при весьма высокой жизненной активности (уровня дыхания в первую очередь). Ситовидные элементы в этом отношении представляют собой уникальное явление среди всех остальных клеток. Впрочем, полное отсутствие ядра отмечено только у цветковых растений, тогда как ситовидные элементы голосеменных и других групп высших растений в большинстве случаев сохраняют ядро, но, как правило, в полуразрушенном состоянии. Такие сморщенные ядра, почти всегда (или даже всегда) лишенные ядрышка, обозначаются как некротические ядра.
Внутренняя организация ситовидных элементов в той мере, как она вскрывается с помощью электронного микроскопа, также не лишена своеобразия. Как правило, в зрелых ситовидных элементах рибосомы полностью отсутствуют, аппарат Гольджи отмечается только на первых стадиях их формирования. Напротив, хорошо развиты митохондрии, пластиды и эндоплазматическая сеть. Вакуоль в этих клетках также отсутствует, что приводит к значительному разжижению цитоплазмы. Сильно развитая эндоплазматическая сеть пронизывает не только цитоплазму ситовидных элементов, но и цитоплазматические тяжи в каналах ситовидных полей. Пластиды также обильны, причем у цветковых растений и некоторых голосеменных можно различать два типа пластид - содержащие белок (Б-тип) и содержащие крахмал (К-тип). Это различие имеет известное таксономическое значение, поскольку большинство двудольных имеет пластиды К-типа, а однодольные - Б-типа. Впрочем, различие это не очень четкое, и нет уверенности, что оно не имеет исключений. Для большинства голосеменных характерно наличие пластид двух типов, тогда как пластиды исследованных в этом отношении папоротников и плаунов не имеют ни белка, ни крахмала.
Крахмал в пластидах К-типа весьма своеобразен и, в частности, не гидролизуется при действии диастазы и альфа-амилазы. Белки в пластидах Б-типа также, по всем данным, отличаются некоторыми особенностями и аналогичны тем белкам, которые скапливаются в полостях ситовидных элементов в виде округлых телец. Эти тельца в недалеком прошлом обозначались как "слизевые тельца", но ныне их принято называть "флоэмным белком" или еще проще - Ф-белок. Этот белок, в зрелых ситовидных элементах теряющий свою округлую форму, устроен своеобразно и, по мнению ряда исследователей, по своей структуре несколько напоминает белок мышц животных. В связи с этим высказывается предположение, что до сих пор еще загадочный механизм движения вещества по ситовидным элементам связан с сократительными реакциями этого Ф-белка, который тем самым гонит вещества вдоль по ситовидной клетке.
Отсутствие или полная некротизация ядра в ситовидных элементах находится в разительном противоречии с интенсивной физиологической активностью этих клеток. Высказывалось предположение, что ситовидные трубки выполняют свои функции лишь в молодом возрасте, пока ядро еще не успело разрушиться. Действительно, у многих растений ситовидные элементы функционируют относительно короткий срок. Все же этот срок редко бывает меньше одного года, а у некоторых растений (особенно у лишенных способности к вторичному росту) продолжительность работы ситовидных элементов составляет многие годы. Поэтому широко распространена точка зрения, по которой управление всей жизнедеятельностью безъядерных ситовидных элементов возложено на ядра соседних клеток.
Действительно, уже у голосеменных можно выявить в непосредственном соседстве с ситовидными элементами особые клетки (альбуминовые клетки, или клетки Страсбургера), характеризующиеся хорошо выраженным ядром. У цветковых растений клетки, прилегающие к ситовидным трубкам, явным образом связаны с ними в своем образовании, являясь результатом последнего митоза материнской клетки. Таким образом, эти клетки являются сестринскими клетками ситовидной трубки. Их постоянное присутствие около ситовидной трубки привело к тому, что их обозначают как "клетки-спутницы" (или более часто, но вряд ли грамматически правильно, как "клетки-спутники"). Любопытной особенностью клеток-спутниц является заторможенность работы их пластидного аппарата, который начинает синтезировать крахмал только после того, как прилегающая к клетке-спутнице ситовидная трубка дегенерирует.
Предположение о своего рода "дистанционном управлении" со стороны ядра соседней с ситовидным элементом клетки всей деятельностью этого последнего широко распространено, хотя, надо признать, экспериментального подтверждения этому нет. Скорее всего эти взгляды основаны на весьма обоснованном убеждении большинства биологов о решающем значении для жизни клетки клеточного ядра.
Мы уже указывали, что ситовидные элементы у разных растений имеют совершенно различный срок активной жизни. У большинства растений, способных ко вторичному росту, ситовидные элементы работают всего одну вегетацию и на следующий год заменяются новообразованными элементами. После этого они или полностью разрушаются или же оболочки их утолщаются (но, как правило, остаются целлюлозными) и эти клетки начинают играть механическую роль. Очень часто полости их целиком исчезают, оболочки соседних клеток смыкаются в однородную массу и возникает своеобразная механическая ткань с характерным перламутровым блеском целлюлозных оболочек, известная под названием "рогового луба".
Исследование структуры флоэмы в целом и особенно ситовидных трубок и в оптическом, и в электронном микроскопах представляет некоторые технические трудности, почему в этой области до сих пор и не накоплен столь обширный сравнительно-анатомический материал, как о строении древесины. Суммируя имеющиеся материалы, можно отметить прежде всего, что для всех цветковых растений, как мы уже указывали, характерно появление ситовидных пластинок, т. е. собрание отдельных групп каналов (отдельных ситовидных полей) в некий, достаточно четко ограниченный участок - ситовидную пластинку. Эти ситовидные пластинки могут состоять из большого числа (10 и более) отдельных ситовидных полей, располагающихся на боковых стенках. Ситовидные трубки при этом длинные, и их боковые стенки только незначительно скошены. Такой тип ситовидной трубки может рассматриваться как наиболее примитивный среди цветковых растений, что подтверждается его сходством с устройством ситовидных элементов голосеменных и приуроченностью к семействам цветковых растений, рассматриваемых как примитивные на других основаниях.
Промежуточным типом ситовидных трубок являются менее длинные ситовидные элементы. У них ситовидные пластинки с небольшим числом (до 10) ситовидных полей располагаются на поперечных стенках (еще несколько скошенных). Наконец, вершиной эволюции ситовидных трубок у двудольных являются относительно короткие клетки с поперечной (или несколько скошенной) стенкой, на которой располагается ситовидная пластинка, где уже невозможно выделить отдельные ситовидные поля.
Клетки-спутницы, по-видимому, претерпевают также некоторую эволюцию. Причем, если сопоставлять их структуру с организацией ситовидных трубок (по описанному только что ряду), то оказывается, что наиболее примитивной структурой является клетка-спутница, возникающая в результате одного митотического деления вместе со своей сестринской клеткой, превращающейся в ситовидную трубку. Последующая эволюция привела к появлению уже нескольких клеток-спутниц, причем исходная (материнская) клетка-спутницу могла делиться и поперечными и продольными делениями, так что в результате возникает целая гирлянда клеток, окружающих ситовидную трубку (клеток, которые приходятся ей, так сказать, "племянницами" и "внучатыми племянницами"). Такое количество клеток, ядра которых, по изложенной выше концепции, должны управлять активностью одной-единственной ситовидной трубки, несколько противоречит простой человеческой мудрости о "семи няньках, у которых дитя без глазу", но вполне вероятно, что мудрость природы в некоторых случаях решительно отличается от нашей.
Кроме покровной и проводящей систем тканей, у высших растений различают еще систему основных тканей, к которой принадлежат все клетки, располагающиеся между эпидермой и проводящими тканями, а также клетки, составляющие сердцевину - массив клеток, расположенный в центре стебля и обычно отсутствующий в корне.
В основной ткани различают две самые крупные группы клеток. К одной группе относятся клетки фотосинтезирующие, содержащие, следовательно, зеленые пластиды (хлоропласты) и сконцентрированные в основном в листьях или в самых молодых частях стебля (особенно у травянистых растений). Это хлоренхима, что в переводе с греческого означает всего лишь "зеленая ткань". К другой группе относят клетки, снабженные бесцветными пластидами - лейкопластами, которые специализированы на превращении и хранении запасных веществ, накопляемых в растении в течение вегетации и используемых в подходящий момент для построения новых частей или образования семян и плодов. Клубень картофеля является характернейшим примером скопления такой "запасающей ткани".
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'