Семейство ореховые (Juglandaceae)
Семейство ореховых включает 7 родов и около 60 видов, широко распространенных в умеренных и субтропических областях северного полушария (карта 16). Многие ореховые произрастают и в тропиках, но главным образом в горах. В южном полушарии встречаются виды только двух родов - ореха и энгельхардии.
Ореховые - деревья (редко кустарники), обычно листопадные, с тонкими ароматическими перистосложными листьями без прилистников. Лишь у видов энгельхардии листья кожистые, сохраняющиеся на дереве большую часть года и опадающие в сухое время.
Древесина плотная, с четкими годичными кольцами (за исключением энгельхардии и южноамериканских видов ореха). У двух родов (ореха и птерокарии) отмечена перегородчатость (септированность) сердцевины в молодых ветвях, которую обычно рассматривают как следствие быстрого роста листьев.
Цветки ореховых однополые, некрупные и невзрачные, как правило, собраны в однополые соцветия (женские цветки иногда одиночные). Околоцветник, если имеется, простой, четырехчленный. Цветки обычно однодомные (двудомными иногда бывают у энгельхардии), расположенные в пазухах прицветников, большей частью цельных (у энгельхардии, ореомуннеи и альфарои трехлопастных). Каждый цветок расценивается, впрочем, как редуцированный до одного цветка трехцветковый дихазий (типа дихазия роиптелеи, но однополый).
Мужские цветки почти у всех видов зигоморфные, с двумя прицветничками, со свободными тычинками (которых может быть от 2 до 105) на коротких нитях; пыльники двугнездные. .а Женские цветки актиноморфные, с двумя прицветничками (иногда отсутствующими) и син-карпным гинецеем из двух плодолистиков (изредка бывает 3-4 плодолистика); рыльце крупное, обычно двулопастное.
Плод костянковидный (за одним исключением), крылатый или бескрылый (рис. 176). Семя крупное, без эндосперма, разделенное обычно на 2 лопасти (иногда на 4 и даже на 8 лопастей).
Основное хромосомное число равно 16. Растения обычно диплоидные (2n = 32), реже тетраплоидные ( 2n = 64); отмечены аномальные числа хромосом.
Значение ореховых в жизни человека велико и многогранно. Древесину практически всех видов применяют в строительстве, но мировая известность - лишь у древесины ореха. Плоды ореха и карий широко используют как пищевой продукт. Кору и оболочку плодов применяют для изготовления красителей. Листья имеют лекарственное значение. По крайней мере у трех родов ореховых листья содержат яд, парализующий рыб. Некоторые индейские племена в Мексике для оцепенения рыб раздавливают молодые листья пекана и ореха наскального (Juglans rupestris) и опускают их пучками в воду. На Суматре местное население для этой же цели использует энгелъхардию Роксбро (Engelhardia roxburghiana).
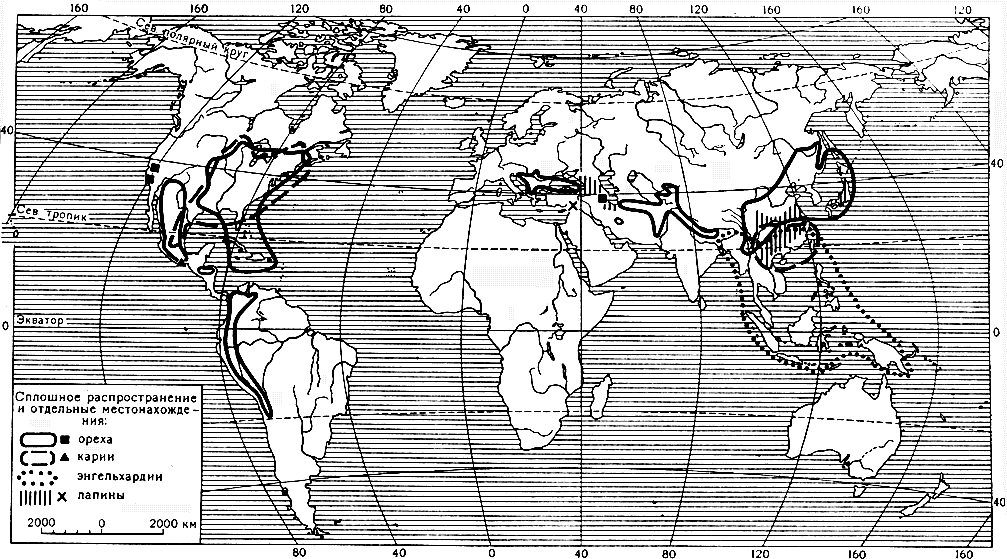
Карта 16. Ареалы родов семейства ореховых
Наиболее известным родом семейства является орех (Juglans), в составе которого, вероятно, насчитывается не более 20 видов (не считая подвидов, которые некоторыми ботаниками возводятся в ранг видов). Самый знаменитый из них - орех грецкий, или царский (J. regia). Научное латинское название было дано этому растению Карлом Линнеем, использовавшим древнеримские его обозначения. Римляне называли плод ореха basilicon (царский), а также juglans, что представляет собой народное сокращение выражения Jovis glans, т. е. желудь Юпитера (Jovis - родительный падеж от Juppiter), иными словами, божественный желудь обладающий в противовес обычному желудю превосходным вкусом.
Чрезвычайно высоко ценимая еще с древности древесина ореха грецкого (легко обрабатываемая, плотная, прочная, не дающая трещин, не меняющая объема под действием тепла и очень красиво окрашенная - от светло-серой до темно-коричневой), известная во многих странах под именем "орех", до сих пор употребляется для изготовления дорогой мебели, ружейных лож и пистолетных рукоятей (преимущественно спортивных моделей оружия), различных поделок. Особую ценность имеют так называемые капы - наплывы при основании стволов, достигающие колоссальных размеров и массы (до 1 т). Популярность древесины ореха столь велика, что на мировом рынке продается много ее заменителей (древесина растений, относящихся к 9 другим родам и 7 семействам).
Грецкие "орехи" и в наши дни ценятся как прекрасный пищевой продукт, употребляемый в сыром виде и в различных кондитерских изделиях. В "Толковом словаре" В. И. Даля "орех" определен как "древесный плод в твердой, крепкой скорлупе". В качестве примеров приведены орех грецкий и орешник, или лещина (Corylus). Однако такое обиходное представление о плоде типа орех не совпадает с ботаническим (нерастрескивающийся сухой односемянный плод). Из двух примеров В. И. Даля лишь плод лещины признается оотаниками настоящим орехом. Путаница происходит из-за того, что плоды ореха грецкого обычно поступают в продажу без внешней мягкой оболочки, состоящей, в понимании ботаника, из двух слоев - наружного тонкого экзокарпия и мягкого промежуточного мезокарпия. Твердая скорлупа грецких "орехов" является внутренним слоем плода - эндокарпием, в данном случае косточкой. Поэтому и сам плод ореха грецкого ботаники называют костянковидным или ложной костянкой (он не является настоящей костянкой; типичные сочные костянки - всем известные сливы, вишни, персики), а по более точной терминологии - нижней (так как развивается из нижней завязи в противовес типичной костянке) сухой синкарпной костянкой. Внутри косточки находится "ядро" - одно семя, лишенное эндосперма, с двумя своеобразными крупными морщинистыми семядолями, разделенными каждая на две лопасти. Из семян добывают масло, имеющее, кроме пищевого, техническое применение (например, в масляной живописи, в полиграфии). Из внешней оболочки плода получают очень стойкие краски (черную и коричневую).

Рис. 176. Плоды ореховых: 1 - орех серый (Juglans cinerea) (а - экзокарпий и мезокарпий, - на рисунке частично убраны, б - ребристый эндокарпий); 2 - пекан (Carya illinoensis) (a - экзокарпий и мезокарпий, б - эндокарпий); 3 - кария китайская (Сагуа sinensis); 4 - птерокария палиурусовидная (Pterocarya paliurus); 5 - лапина кавказская (Pterocarya fraxinifolia); 6 - энгельхардия Вал-лиха (Engelhardia wallichiana); 7 - платикария шишконосная (Platycarya strobilacea), 1 - 6 - увел. 0,8; 7 - увел. 2,4
По русскому названию вида можно думать, что родиной ореха грецкого является Греция. И он действительно в диком состоянии растет в Греции. Однако у ботаников имеются серьезные основания считать, что орех грецкий лишь одичал здесь много веков назад. Интересно, что, кроме чисто ботанических аргументов, к решению этого вопроса можно привлечь также данные из области языкознания!
В самом деле, попробуем установить, как возникло русское словосочетание "грецкий орех"? Сама форма слова "грецкий" подсказывает нам, что оно является древним в русском языке: без слова "орех" оно не употребляется в наши дни в живой речи. В книжном же языке его изредка можно встретить как архаизм и обязательно в сочетании с другими словами, например "грецкая губка" (морская губка), "грецкое вино", обозначающими предметы былой торговли иностранных купцов на Руси. Словом "Греки" наши предки обозначали Византию ("путь из Варяг в Греки"), которая прекратила свое существование под ударами турок-османов в 1453 г. Следовательно, слово "грецкий" применительно к византийскому импорту могло возникнуть на Руси до последнего события. Но заглянем еще глубже в историю. Оказывается, что и знаменитый римский ученый Марк Теренций Варрон (I в до н. э.) в труде "О сельском хозяйстве" называл орех греческим (juglans mix graeca). В Рим грецкий орех попал, следовательно, из Эллады. Обратившись к самому раннему в истории человечества научному труду по ботанике, автором которого был древнегреческий ученый Теофраст (IV - III вв. до н. э.), мы увидим, что он называл это растение персидским орехом. Но в XIX в., когда начался расцвет палеоботаники, остатки орехов (плоды и листья) были обнаружены в третичных и четвертичных отложениях Европы. Это показалось ученым того времени несомненным доказательством непрерывного существования ореха грецкого в Европе с третичного времени. Лишь более точными исследованиями, проведенными в наши дни, было установлено, что эти ископаемые орехи родственны не ореху грецкому, а другим видам рода, сохранившимся до наших дней лишь в Северной Америке.
Учитывая свидетельство Теофраста и современные данные о распространении ореха грецкого, можно считать областями его, безусловно, естественного распространения Южный Казахстан, Среднюю Азию, Иран, Афганистан,; западные части Гималаев и Тибета, юго-восток Закавказья (Талыш). В других частях Закавказья, например в Западной Грузии, огромные леса из ореха грецкого справедливо расцениваются как разросшиеся древние сады, заброшенные еще во время грузино-персидских и грузино-турецких войн. Путешествовавший в 1924 г. по Афганистану замечательный русский генетик, растениевод и ботаник Н. И. Вавилов писал, что Афганистан в целом входит в общий ареал дикого ореха грецкого, но в ряде замкнутых районов, например в Кафиристане, имеются следы древней самостоятельной культуры этого растения. Также и в Малой Азии и в Европе дикорастущие орехи грецкие следует считать потомками одичавших культурных растений. О том, что в Европе действительно было так, имеется и лингвистическое свидетельство.
Если мы нанесем на карту названия ореха грецкого на всех европейских языках, то обнаружим, что у романских народов, внутри бывшей Римской империи, название ореха грецкого преобразовалось просто в "орех" (нук по-молдавски, nuc по-румынски, посе по-итальянски, nogal по-испански, словом, все производные от латинского mix). На окраинах же Римской империи и у ее германских и славянских соседей орех грецкий получил название "волошский орех": по-чешски, orzeeh по-польски, Walnuss по-немецки, по-датски, по-английски и даже у украинцев - горiх волоський. Волб-хами же называли в прежнее время чужеземцев, римлян (древневерхненемецкое Walh). И сейчас еще так называют румын, обитателей Валахии. Ясно, что в течение многих веков орех грецкий, разводимый почти повсеместно на территории Римской империи, ввозился в более северные страны и там делались попытки вводить его в культуру. Ведь в наши дни удается выращивать его и в Ленинграде, и в Норвегии (где имеются плодоносящие экземпляры).
Орех грецкий разводится теперь почти во всех странах мира и во многих местах дичает. Он вполне натурализовался, например, в Северной Америке, где, как установлено, его плоды запасает впрок индейский медведь (Ur-sus torquatus), содействуя тем самым распространению этого растения. Разносят плоды ореха и некоторые птицы. Белки срывают плоды с ветвей, закапывают в землю, тщательно маскируя. Они используют запасы некоторое время спустя, когда внешний слой плода разрушается. Разумеется, часть спрятанных плодов остается неотысканной, они прорастают и дают начало новым особям.
В результате многовековой культуры выведено огромное количество сортов ореха грецкого, в том числе мягкоскорлупый, мамонт (крупные плоды), бомба (очень крупные плоды, тонкая скорлупа), рано созревающий (цветет уже в трехлетнем возрасте, тогда как дикие экземпляры - лишь с 10 лет), специальные сорта для получения масла и т. д.
Плоды и древесина ореха грецкого известны каждому, но его цветками мало кто интересуется. Более того, в Средней Азии многие старики, всю жизнь наслаждавшиеся тенью этих высоких, с обширной кроной деревьев, считают, что орех грецкий не цветет вовсе ("Умрет тот, кто увидит цветок ореха", - говорят они). Действительно, на большой высоте невзрачные цветки ореха грецкого малозаметны, да и просто не считаются за цветки.
Мужские цветки собраны в зеленые, свисающие в период цветения сережки, длина которых доходит до 12 см. Сережки закладываются летом на молодых побегах, к осени они представляют собой крупные конические почки, которые в таком состоянии перезимовывают, а в апреле - мае распускаются одновременно с листьями, и происходит быстрый рост соцветия. Каждый цветок располагается в пазухе прицветника. Начало формирования цветка относится к июню предыдущего года. Первыми образуются 2 меристематических бугра (будущие прицветнички), следом за ними еще 4 (листочки простого околоцветника, срастающиеся впоследствии основаниями). Одновременно в неопределенном количестве возникают бугорки - будущие тычинки. Из тычиночных бугров ранней весной формируются тычинки на коротких нитях. В верхних цветках соцветия их по 6 - 8, в нижних - по 20 - 30.
Опыление происходит с помощью ветра. Пчелы посещают преимущественно мужские цветки (ради пыльцы), поэтому их участие в опылении не может быть существенным. Перекрестное опыление обеспечивается разновременностью развития мужских и женских цветков (разница в сроках цветения - 15 суток) на одном растении (протогиния, реже протандрия). Пыльцевые зерна (округлой формы, 3 - 4-поровые до многопоровых) ореха грецкого быстро теряют способность к прорастанию (через 2 - 3 суток). При большой густоте насаждений это приводит к перенасыщению воздуха стерильной пыльцой, которая инактивирует рыльцевые поверхности, и процент оплодотворенных женских цветков резко снижается. Видимо, из-за этого наиболее урожайны одиноко стоящие деревья (некоторые гигантские экземпляры дают до 50 тыс. плодов в год), у которых преобладают женские цветки. Плантации ореха грецкого приходится поэтому закладывать с учетом расстояний между взрослыми особями.
Женские цветки одиночные или собраны по 2 - 4 в верхней части побегов. Каждый цветок расположен в пазухе прицветника, снабжен двумя прицветничками и простым околоцветником, состоящим из четырех сросшихся основаниями листочков.
При вторичном цветении (осенью) иногда образуются обоеполые цветки. Подобные аномалии вообще довольно нередки у культурных и одичавших растений. В Киргизии и Таджикистане ботаник Т. Дускабилов обнаружил также экземпляры, дающие обоеполые цветки весной, при первичном цветении. Эти растения к тому же необычайно рано - уже на первых годах жизни - дают цветки и плодоносят.
Описаны также аномальные случаи образования обоеполых соцветий (в нижней части - женские, в верхней - мужские цветки). Это было замечено, например, во Франции у культурных сортов ореха (на привитых экземплярах).
Орех грецкий является лесообразующим деревом. Леса из ореха грецкого встречаются даже в районах, где это растение заведомо не аборигенное.
В Средней Азии, где первичность существования ореха грецкого несомненна, ореховые леса и редколесья (с участием яблони, груши и арчи) расположены на высоте 1000 - 2000 м, а отдельные растения спускаются и гораздо ниже, но вверх не заходят выше 2300 м. Осенью, обычно в сентябре, после обильных дождей и первых заморозков косточки ("орехи" в житейском смысле) начинают выпадать из наружной оболочки. Местное население интенсивно собирает грецкие "орехи" и в лесах, несмотря на обилие культурных экземпляров в поселках.
Сложные листья ореха грецкого состоят из 5 - 7 (редко из 9 или 3), как правило, цельно-крайних листочков (у остальных видов ореха листочки зубчатые). Обычно верхние листочки крупнее нижних, а для подвида ореха грецкого обманчивого (J. regia subsp. fallax), обитающего в Средней Азии (по мнению ряда ботаников - вплоть до Индии), характерны очень крупные конечные листочки (ср. рис. 10).
Орех грецкий - единственный современный представитель секции югланс (Juglans) рода орех, если, конечно, не принимать подвиды, подобные только что упомянутому, за отдельные виды.
В пределы СССР заходят еще два вида - орех маньчжурский (J. mandshurica) и орех айлантолистный (J. ailaiithifolia, табл. 47), чаще называемый орехом Зиболъда (J. sieboldiana). Орех маньчжурский обитает в Хабаровском и Приморском краях нашей страны, а также в Северном Китае и северной части полуострова Корея. Это наиболее далеко заходящий на север вид ореха (до 51° северной широты). Его древесина пригодна для всевозможных столярных работ, но он сильно истреблен и на рубку требуется специальное разрешение. Плоды ореха маньчжурского обладают очень твердой косточкой, семя занимает (сравнительно с плодами ореха грецкого) меньший объем, а вкусовые качества невысоки. Тем не менее незрелые плоды употребляют для изготовления варенья (подобно тому как это делается из плодов ореха грецкого, но после длительного вымачивания). Известно, что плодами ореха маньчжурского питаются кабаны и некоторые другие животные. Листья у ореха маньчжурского крупнее, чем у ореха грецкого, состоят из 9 - 19 листочков.
В приатлантических штатах США обитает орех серый (J. cinerea). Прежде ареал этого вида был значительно шире - его ископаемые остатки обнаружены в четвертичных отложениях Евразии. Многие третичные виды ореха, обнаруженные в Северной Америке, Европе и Северной Азии, родственны именно ореху серому. Орех серый имеет промышленное (древесина, желтая и оранжевая краски из внешней оболочки плода) и пищевое значение. Весьма декоративен, разводится, помимо США, в Европе. Этот вид вместе с орехом маньчжурским, орехом айлантолистным и орехом катайским (J. cathayensis) составляют секцию кардиокарион (Cardiocary-оп) рода орех.
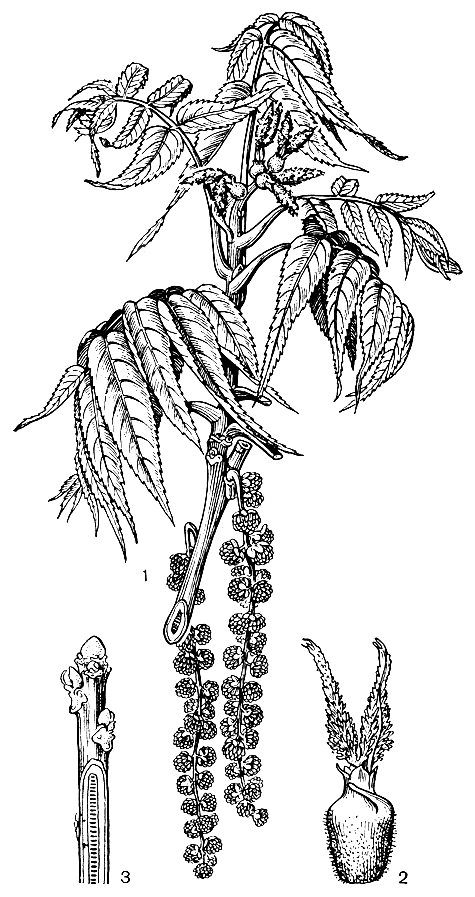
Рис. 177. Орех черный (Juglans nigra): 1 - ветвь с молодыми листьями, женскими цветками (в верхней части) и мужскими соцветиями (в нижней части); 2 - женский цветок; з - молодой побег (на срезанной части показана септированная сердцевина)
Другой обитатель восточной части Северной Америки, орех черный (J. nigra, рис. 177), пищевое, техническое и декоративное растение, вместе с орехом калифорнийским (J. californica), мексиканскими, вест-индскими и пятью южноамериканскими видами объединены в секцию ри-зокарион (Rhysocaryon). Все южноамериканские орехи растут в горах (орех неотропический - J. neotropica - в Эквадоре до высоты 3000 м). Ареал ореха южного (J. australis) достигает севера Аргентины.
Линней включил в род орех, кроме ореха грецкого, также американские виды. Уже в конце XVIII в. ботаники начали различать в линнеевском роде орех 2 группы видов - собственно орехи и хикори (индейское название), а в 1818 г. хикори были описаны как особый род кария (Саrуа). Слово karya означало у древних греков "орешник"; чаще всего под этим понимался орех грецкий. Поначалу были известны только американские виды карий, из которых наиболее важным является пекан,или иллинойский хикори (С. illinoensis, рис. 178, 179, табл. 48), имеющий в Северной Америке то же значение,: что и орех грецкий в Евразии. Карий отличаются от орехов прежде всего строением плодов (рис. 176, 179). Внешняя оболочка у карий растрескивающаяся обычно на 4 створки, косточка гладкая, а у видов ореха, как каждый может убедиться, взяв в руки нерасколотый грецкий "орех", - нерегулярно-бороздчатая. Мужские соцветия карий собраны в пучки по 3 - 8 у разных видов, а у ореха - одиночные или собраны по 2. Околоцветник у карий отсутствует. Те 4 листочка, которые могут быть обнаружены в женском цветке вокруг гинецея, не являются настоящим околоцветником. Это прицветники.
Род кария содержит 16 (или 18) видов. Почти все карий - крупные деревья, за исключением карий флоридской (С. floridana), кустарникового хикори. Этот вид относится к целиком американской секции кария (Сагуа), в которую входит еще 6 видов, распространенных в восточных частях Северной и Центральной Америки. Один из них, кария опушенная (С. tomentosa), - типично лесной вид. Она распространена от Великих озер до Мексиканского залива и от востока Техаса до берегов Атлантики. Карию опушенную называют фальшивым орехом (mockernut), так как ее крупный плод обладает очень толстой наружной оболочкой, внутри которой содержится неожиданно маленькое семя (рис. 179).
Плоды американских карий, в том числе и тех, которые человек не использует, поедаются утками, индюками, куропатками, рябчиками, фазанами, лисами, кроликами, бурундуками, белками, вирджинским оленем.
Секция апокария (Apocarya) содержит 8 видов, из них 4 вида распространены в восточной части Северной Америки. Сюда относится и знаменитый пекан. Он широко разводится ради съедобных плодов и получаемого из них масла. Ценится и его древесина. Естественной частью ареала пекана считается долина Миссисипи (от юга Индианы, севера Иллинойса и юго-востока Айовы до Алабамы, Миссисипи, Луизианы и востока Техаса). В диком состоянии встречается также в Мексике. Но точно определить область его первоначального распространения практически невозможно, так как он искусственно разводился еще древними индейцами (в Луизиане, Миссисипи, Техасе) и во многих местах одичал.

Рис. 178. Женские цветки пекана (Garya illinoensis) в Сухумском ботаническом саду
Новая жизнь пекана в культуре началась, впрочем, совсем недавно, в середине прошлого века, но была столь бурной, что к настоящему времени только в США насчитывают не менее 100 культурных форм этого вида. Наиболее интенсивно пекан разводится как пищевое и декоративное растение в Джорджии и Флориде.
Известен в культуре во многих странах мира: в Западной Европе (Франция, Испания), в СССР (Черноморское побережье от Сочи до Батуми, Крым, Ленкорань, Средняя Азия), в Турции, в Австралии. Как и все ореховые, пекан является ветроопыляемым растением.
Длинные свисающие мужские сережки пекана расположены по 3 в пучке, пучок - на укороченном побеге. По 2 - 4 таких укороченных побега вырастает около листовых рубцов на прошлогоднем побеге, а всего прошлогодний побег несет до 5 - 7 групп укороченных побегов. Если учесть, что в одной сережке может быть 100 - 150 цветков, а в цветке 4 - 7 тычинок, получается гигантское количество пыльников на одной годичной ветви - около 400 тыс. Одно крупное дерево пекана дает многие миллиарды пыльцевых зерен. А пеканы обладают удивительной особенностью - пыльники при благоприятных условиях (в теплую погоду при средней влажности воздуха) раскрываются очень быстро и почти одновременно. Пыльца при этом молниеносно высыпается. Весь процесс происходит в течение двух-трех послеполуденных часов одним - тремя импульсами. Над деревом внезапно, как бы в результате взрыва, появляется заметное глазу облачко пыльцы, которая при наличии ветра сразу относится в сторону.

Рис. 179. Плоды карий: вверху - пекан (Сагуа illinoensis); внизу - кария опушенная (С. tomentosa). Батумский ботанический сад АН Груз. ССР
У карий происходит перекрестное опыление и самоопыление, но при самоопылении плоды7] завязавшись, опадают в большом количестве недозрелыми. Интересно, что и изобилие пыльцы, и слишком малое ее количество может сказаться отрицательно на оплодотворении.
Женские соцветия пекана содержат от 3 до 11 цветков (рис. 178). Ко времени цветения обычно верхушечные цветки остаются недоразвитыми и опадают неоплодотворенными. Женские цветки пекана не имеют привычной нам стадии бутона: в начальной фазе созревания цветка наблюдается отгибание верхних частей сросшихся между собой прицветников, рыльце, представленное в этот момент едва заметным темно-зеленым бугорком, начинает расти и через 5-7 суток достигает полного развития, изменяя при этом свой цвет на светло-зеленый. Две крупные лопасти рыльца покрыты в зрелом состоянии многочисленными сосочками, выделяющими в течение нескольких дней значительное количество опылительной жидкости, так что рыльца становятся липкими. При избыточном или малом количестве пыльцы рыльца темнеют и высыхают, и неоплодотворенный цветок опадает.
Пекану свойственна как протандрия, так и протогиния. У одного растения от сезона к сезону тип дихогамии может меняться, что в какой-то степени зависит и от погодных условий до цветения.
Анемофилия пекана проявляется и в динамике всего процесса цветения и опыления (взрывчатость, порционность цветения, приуроченность его к определенному времени суток) - это называют динамической анемофилией, и в специальных устройствах его цветков, т. е. в структурной анемофилии: раздельнополость цветков, их невзрачность (отсутствие яркой окраски, небольшие размеры - кроме рылец!), отсутствие настоящего околоцветника и нектарников, сыпучесть и легкость пыльцевых зерен, расположение женских цветков на верхушках побегов (тем самым - в наружной части кроны), большая "рабочая" поверхность и долговечность рылец.
Пеканы (как, впрочем, и некоторые орехи, например орех черный) во время цветения являются причиной аллергических пыльцевых заболеваний.
Пятый американский член секции апокария, кария Палмера (С. palmeri) - эндемик Мексики. Она родственна горькому хикори - карий сердцевидной (С. cordiformis), обитателю Юго-Восточной Канады и восточной части США. Кария сердцевидная, растущая и в явных субтропиках на севере Флориды, и в умеренной зоне, имеет наибольший среди карий естественный ареал.
Эти два американских вида более других родственны двум азиатским видам карий, карий катайской (С. cathayensis) и карий тонкинской (С. tonkinensis), входящим в секцию апокария. Такое удивительное разъединение (Северная Америка и Южная Азия) близкородственных видов, относящихся не только к одному роду, но даже к одной и той же секции рода, долгое время волновало ботаников. Знаменитый американский ботаник-дендролог Ч. С. Сарджент, описавший в 1916 г. карию катайскую, был озабочен прежде всего поиском ее родства с американскими кариями. Он посчитал тогда наиболее близким видом карию мускатниковидную (С. myristiciformis) из секции апокария. Но именно из состава этого вида другой американский ботаник, У. Маннинг, выделил в 1949 г. карию Палмера. Следовательно, Ч. С. Сарджент был прав, ибо из всех американских видов наиболее близка азиатским как раз кария Палмера.
Как же могло случиться, что близкие родственники оказались столь разобщенными территориально? Разобраться в этом помогают данные палеоботаники. В третичное время ареал секции апокария был значительно шире. На севере Евразии, от Франции и Италии до Абхазии, в течение примерно 15 млн. лет существовала кария мелкозубчатая (С. denticulata). Она родственна карий тонкинской. А к северу от нынешних мест обитания карий катайской в третичное время существовал близкий ей вид, ныне вымерший. В других же частях Евразии почти повсеместно в третичных лесах встречались виды карий, родственные современным американским (кариям сердцевидной, опушенной и др.). В конце третичного времени, а особенно в четвертичное время в северном полушарии произошло сильное похолодание и ареал рода сильно сократился. Исчезло и соединение Америки с Азией, так называемый Берингийский мост, а с ним утратились и связи между родственниками. Остатки некогда большого рода кария оказались как бы разбросанными по земному шару после всемирной климатической катастрофы.
К третьей секции рода кария - рамфокарии (Rhamphocarya) - относится только кария китайская (С. sinensis, рис. 176), распространенная во Вьетнаме и Китае (Юньнань и Гуйчжоу). Этот вид обладает цельнокрайними листочками, не встречающимися у других карий, и очень крупными плодами (диаметром более 5 см). Он был описан в 1912 г. французским ботаником Л. Додом как первая находка карий в Азии (до этого времени род считался американским). Однако эта публикация была странным образом забыта и по новым находкам карий китайской в 1941 г. два ботаника, француз О. Шевалье и китаец Куан Кэжень, опубликовали каждый отдельно, не зная друг о друге и о статье 1912 г., два новых рода, точнее, два новых родовых названия - аннамокарию (Annamocarуа) и рамфокарию (Rhamphocarya). На сей раз публикации были замечены, ибо наступила эпоха интереса к "живым ископаемым": как раз в это время описывается ископаемый род хвойных - метасеквойя и вскоре его обнаруживают в живом состоянии (подробнее см. "Жизнь растений", т. 4). По аналогии с метасеквойей в аннамокарии (рамфокарии) некоторые ботаники увидели живого представителя ископаемого рода - либо югландикарии (Juglandicarya), либо кариоюгланса (Caryojuglans), незадолго до этого описанных по европейским материалам. А дело в том, что кария китайская (она ведь и была описана как аннамокария и рамфокария) обладает сходством не только с ка-рией, но и с орехом. Начиная с 1948 (когда появилось сравнение с югландикарией и кариоюг-лансом) по 1953 г. (когда появилась обстоятельная статья американского ботаника Ричарда Скотта с опровержением родства карий китайской и ископаемых родов) в ботанической литературе нескольких стран мира шла оживленная дискуссия о "живом ископаемом" - карий китайской.
Китайская флора удивительно богата и все еще не исчерпана. Достаточно сказать, что из семи родов ореховых пять представлены в Китае. Это роды орех, кария, птерокария, плати-кария и энгельхардия. Добавим сюда и необычные виды перечисленных родов, которым иногда придается родовой ранг (аннамокария и цик-локария, о которой речь впереди).
Птерокария (Pterocarya) была третьим по порядку описания в ботанической литературе родом ореховых. Европейцы познакомились с ней в конце XVIII в., после путешествия французского ботаника А. Мишо по Передней Азии. Мишо собрал не только гербарий, но, видимо, привез и зрелые семена. Во всяком случае, первый раз птерокария была описана как орех ясене-листный (Juglans iraxinifolia) в 1797 г. по экземпляру из сада Парижского музея. Лишь в 1824 г. этот вид был описан как отдельный род птерокария. По кавказскому названию вида (лапина, от груз, лапани) весь род птерокария в русской ботанической литературе часто именуют лапиной.
Птерокария ясене листная, или кавказская (Pterocarya iraxinifolia), называемая обычно лапиной, обитает в СССР на Черноморском и Каспийском побережьях Кавказа, в Турции и Иране на южном берегу Каспийского моря (встречается и в понижениях ниже уровня Каспийского моря). Заходит в горы обычно до высоты 600-800 м, например в Алазанской долине по ущельям притоков Алазани. Отдельные экземпляры зарегистрированы на северных склонах Эльбруса и на Большом Кавказе (на высоте до 1200 м). Лапина - дерево высотой до 35 м, с диаметром ствола 40 - 80 см, но иногда и до 2 м. Порой образует и небольшие чистые заросли но обычно растет в смешанных лесах.
Листья лапины, как правило, непарноперистосложные, длиной 9 - 30 см, с 5 - 15 зубчатыми листочками. Цветоносные побеги лапины закладываются в начале июля в пазухах последних или предпоследних на побеге листьев. На верхушке побега возникает, как правило, зачаток женской сережки (иногда же - вегетативная почка), а ниже ее по всей длине побега располагаются почки мужских сережек. В конце октября цветоносный побег достигает в длину 2 - 6 см, все чешуи, в пазухах которых расположены (по одной) мужские сережки, опадают, сережки вытягиваются в длину. Женская верхушечная сережка к этому времени уже развилась и обогнала в росте мужские сережки. С наступлением морозов опадают листья и цветоносный побег перезимовывает. Ранней весной продолжается рост и развитие сережек. Зрелая женская сережка может быть длиной 10 - 14 см, мужские - 8 - 11 см. Цветение происходит в конце марта - апреле. У лапины бывает и одновременное раскрывание мужских и женских цветков, и оба типа дихогамии (протогиния и протандрия). Зрелая женская сережка (отстающая в росте в случае протандрии) содержит до 70 цветков, но полного развития достигают лишь цветки в средней части соцветия.
После опыления и оплодотворения рыльца засыхают и начинается развитие завязи. Вначале происходит увеличение размеров плода, а в августе очень быстро растет семя. Плод лапины сухой, костянковидный, достигает в диаметре 1 см. Экзокарпий кожистый, образуется за счет разрастания наружной части плодолистиков и оснований частей цветочного покрова (прицветника, прицветничков и листочков околоцветника). Мезокарпий в плоде лапины не выражен. Деревянистый эндокарпий составляет основную часть плода по объему и массе. Семя с двумя крупными семядолями, разделенными на 2 лопасти. Плод с двумя крыльями, представляющими собой разрастание экзокарпия.
После оплодотворения женские сережки лапины становятся соплодиями. Плоды в них увеличились в размерах (сравнительно с завязями), обрели крылья, стали неузнаваемы. Свисающие сережки, вначале зеленые, затем желтеющие и буреющие, на всех стадиях развития красивы, особенно красивы они осенью. Сережки придают прекрасному дереву лапины чрезвычайно изящный вид (табл. 47).
В роде птерокария 11 видов (некоторые ботаники признают только 6), распределенных по двум подродам. Подрод птерокария (Pterocarуа) содержит 10 видов. В нем 2 секции. Секция птерокария включает, кроме лапины, птерокарию узкокрылую (P. stenoptera, табл. 47), птерокарию хубэйскую (P. hupehensis), птерокарию пильчатую (P. serrata), которые распространены в Юго-Восточном Китае, и птерокарию тонкинскую (P. tonkinensis), растущую во Вьетнаме и Лаосе. Секция платиптера (Platyptera) также состоит из 5 видов, из них 4 вида сосредоточены в Центральном Китае, пятый - птерокария сумахолистная (P. rhoifolia) - в Японии.
В подрод циклокария (Cyclocarya) входит лишь птерокария палиурусовидиая (P. paliurus) - замечательный вид, описанный русским ботаником А. Ф. Баталиным в 1893 г. В отличие от представителей подрода птерокария этот вид, обитающий в Центральном и Юго-Восточном Китае, обладает плодами не с двумя крыльями, а с одним крылом, которое в виде кожистого диска облекает весь плод. Этот признак послужил, по-видимому, в первую очередь основанием советскому ботанику И. А. Ильинской для выделения в 1953 г. птерокарии палиурусовиднои в отдельный род циклокария (Cyclocarya), что отражено и в его названии (от греч. kyklos - круг, колесо). Имеются и другие отличительные признаки птерокарии палиурусовиднои от видов подрода птерокария: мужские сережки у нее в пучках по 2 - 4 (чаще по 3), мужские цветки актиноморфные (это, пожалуй, не менее существенный признак, чем наличие кругового крыла у плода) и т. д. Тем не менее многие ботаники не признают птерокарию палиурусовидную в качестве особого рода, так как число сходных между двумя подродами птерокарии признаков больше числа отличительных. Наиболее четко это выразил У. Маннинг, показав, что на стадии цветения невозможно выделить подрод циклокария как особый род. Лишь в процессе развития плода возникает признак, кажущийся родовым: циклическое крыло у птерокарии палиурусовиднои в противовес двум крыльям остальных птерокарии (рис. 176).
Геологическая история обоих подродов сопряженна в третичное время они были весьма широко представлены во флорах Евразии от Франции и Италии до Камчатки.
В середине XIX в. познание флоры Китая шло быстрыми темпами. В Лондон, бывший едва ли не центром мировой ботаники того времени, поступали многочисленные коллекции из Китая. В одной из них, присланной английским военным врачом Т. Кантором в 1840 г., была зарегистрирована "пустая шишка неизвестного хвойного". Спустя 4 года от шотландского "охотника за растениями" Роберта Форчуна был получен китайский гербарий, в котором английский ботаник Дж. Линдли обнаружил растение как раз с такими "шишками", оказавшимися соплодиями растения, похожего по листьям на сумах. Крылатые плоды его напоминали внешне и по размеру плоды ольхи (рис. 163, 176). Дж. Линдли назвал растение в 1846 г. в честь коллектора форчунеей китайской (Fortunea chinensis). Однако почти через 20 лет, когда Казимир де Кандолль (внук и сын знаменитых ботаников - Огюста Пирама и Альфонса де Кандоллей) занимался обзором всех известных к тому времени ореховых, он обнаружил, что Дж. Линдли был опережен известными немецкими исследователями японской флоры Ф. Зибольдом и Й. Цуккарини, которые в 1843 г. назвали это растение платикарией шишконосной (Platycarya strobilacea). Под этим именем оно и значится теперь в ботанических руководствах (рис. 180, табл. 48).
Платикария шишконосная - небольшое дерево или кустарник, обитает в горах Юго-Восточного Китая (до высоты 2000 м), на Корейском полуострове и в Японии. В странах Западной Европы культивируется с декоративной целью. В СССР растет в Крыму в Никитском ботаническом саду и в ботанических садах Черноморского побережья Кавказа.
Платикарии очень красивы. Их декоративность еще более возрастает в цветущем состоянии благодаря зеленым шишковидным женским соцветиям, которые на стадии плодоношения становятся деревянистыми и приобретают коричневую окраску. В Китае плоды используют для окрашивания одежд в черный цвет.
Род энгелъхардия (Engelhardia, рис. 181) был описан в 1825 г. Он получил свое имя в честь тогдашнего губернатора Явы, голландца Э. Энгельхарда. В роде энгельхардия 5 видов, распространенных в субтропических и тропических районах Азии, как правило, в горах (энгелъхардия пильчатая - Е. serrata - от уровня моря до высоты 2200 м). Одни виды энгель-хардии двудомны, другие могут быть и однодомными, и двудомными.
Энгельхардии - крупные деревья, высотой до 47 м, с диаметром ствола до 3 м. У многих старых экземпляров имеются досковидные корни высотой до 4 м, выступающие от ствола на 2 - 3 м. Столь крупные деревья, естественно, используются местным населением (для постройки каноэ, жилищ).
Листья энгельхардии спирально расположенные, парноперистосложные, длиной обычно от 3 до 30 см, но не более 40 см; листочки зубчатые или цельные, бумажистые или кожистые. Энгельхардии большую часть жизни покрыты листьями; лишь утратив их на короткое время в конце сухого сезона, они зацветают. Цветки энгельхардии однополые и по типу строения не отличаются от цветков других ореховых. Части цветка обычно остаются при плоде. Трехлопастной прицветник разрастается в большое тонкое крыло, также трехлопастное, а у энгельхардии колосовидной (Е. spicata) развивается и противопоставленная трем другим четвертая лопасть, прикрывающая плод. Этот признак сближает энгельхардию с ореомуннеей, о которой речь впереди.

Рис. 180. Платикария шишконосная (Platycarya strobilacea): 1 - часть побега с женскими соцветиями; 2 - женский цветок; 3 - часть побега с мужскими соцветиями; 4 - мужской цветок
Плод энгельхардии небольшой, округлый, диаметром 5 - 10 мм (редко 20 мм), вместе с крылом напоминает плод граба. Главная лопасть плодов достигает в длину (вместе с плодом) 7,5 см при ширине до 2 см (рис. 176, 181). Знаменитый французский палеоботаник Адольф Броньяр первым обнаружил ископаемые остатки энгельхардии в Европе в 1828 г. Это был отпечаток плода с трехлопастным крылом. Но Броньяр еще не знал о существовании современного рода с подобными плодами и назвал свою находку грабом крупнокрылым (Carpinus macroptera). Впоследствии выяснилось, что этот вымерший 15 - 25 млн. лет назад вид был широко распространен в Европе и Закавказье. Он называется теперь энгелъхардией крупно-крылой (Engelhardia macroptera), хотя среди энгельхардии его плоды не выглядят такими уж крупными.
Казалось бы, маленькие плоды с большим крылом должны обязательно распространяться ветром. Так оно и происходит в большинстве случаев, однако иногда соплодия (а они бывают довольно длинными, у энгельхардии колосовидной - до 60 см) с крылатыми плодами опадают целиком, не распадаясь, и лежат под материнским деревом.
Геологическая история энгельхардии, как и других ореховых, указывает на ее былое распространение, с начала третичного времени,, в ныне умеренных областях Евразии, от Великобритании до Японии (климат этих мест в третичное время был существенно теплее современного). Ископаемая энгельхардия отмечена и в Северной Америке. Но здесь для более полной картины мы должны обратиться еще к одному роду ореховых - ореомуннее (Oreomunnea, рис. 182).
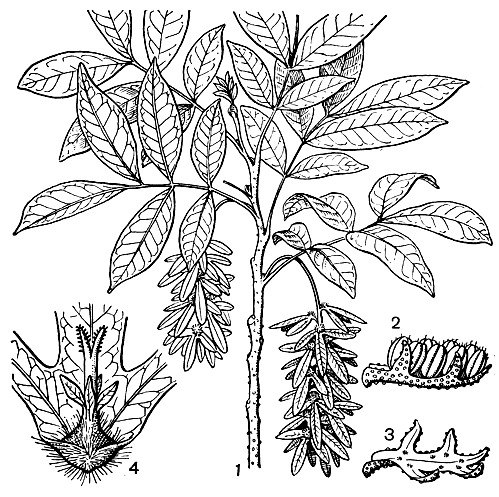
Рис. 181. Энгельхардня жесткая (Engelhardia rigida): 1 - ветвь с листьями и соплодиями; 2 - мужской цветок; 3 - мужской цветок с удаленными тычинками (видны 2 лопасти околоцветника и 3 лопасти прицветника); 4 - плод с остающимися частями околоцветника
Ореомуннея была обнаружена и описана датским ботаником А. С. Эрстедом в 1856 г. Он назвал ее по имени своего помощника в путешествии по Вест-Индии Франсиско М. Ореамуно (впоследствии государственного деятеля Коста-Рики). Эрстед имел в своем распоряжении только странный плод, найденный им на земле во влажных лесах Атлантического побережья Коста-Рики. Самого растения он так и не увидел. Плод этот, как и у энгельхардии, был с трехлопастным крылом (и противопоставленной ему четвертой лопастью), но кожистым и гораздо более крупным. По очертанию он очень напоминал плод энгельхардии, что и побудило Казимира де Кандолля в 1862 г. присоединить этот вид, ореомуннею крылоплодную (О. pterocarpa),; к роду энгельхардия. Однако находка в 1914 г. деревьев ореомуннеи крылоплодной (они оказались весьма высокими - до 48 м при толщине ствола до 70 см) позволила ботаникам восстановить род ореомуннея. Более тщательное исследование плодов ореомуннеи крылоплодной показало, что жилкование лопастей крыла у этого вида пальчатого типа. Кроме средней жилки, которая проходит к верхушке лопасти и, не доходя до нее, разветвляется на более тонкие жилки, из основания в лопасть входят параллельно средней две сильные жилки, анастомозирующие со средней (рис. 182). У энгельхардии же жилкование лопастей более обычного, перистого типа. Имеются отличия и в жилковании (последнего порядка) листочков, в количестве перегородок в плоде, в расположении семядолей, в типе прорастания семян (подземном у ореомуннеи и надземном у энгельхардии) и т. д.
Второй вид ореомуннеи был открыт в 1927 г. в гербарии американским ботаником П. Станд-ли. Оказалось, что еще в 1891 г. некий любитель ботаники - инженер X. Ровироза собрал в Мексике и передал Филадельфийской академии естественных наук (США) гербарий, собранный с дерева, оказавшегося ореомуннеей мексиканской (О. mexicana). Это растение обитает во влажных местах в горах на высоте 900 - 1000 м. Ореомуннея мексиканская имеет признак, отсутствующий у всех остальных ореховых, - ушки у основания листочков. К тому же эти ушки завернуты навстречу друг другу таким образом, что образуют бокаловидную структуру. Листочки ореомуннеи крылоплодной лишены ушек, однако их края у основания тоже несколько завернуты внутрь. Различаются виды ореомуннеи и по величине плодов - у вида, открытого Эрстедом, крыло плода длиной 8 - 15 см (при диаметре собственно плода 10 - 12 мм), а у ореомуннеи мексиканской крыло не достигает в длину и 5 см (диаметр собственно плода 6 - 7 мм). Оба вида ореомуннеи являются эмергентами в тропических дождевых лесах Южной Мексики и Центральной Америки.
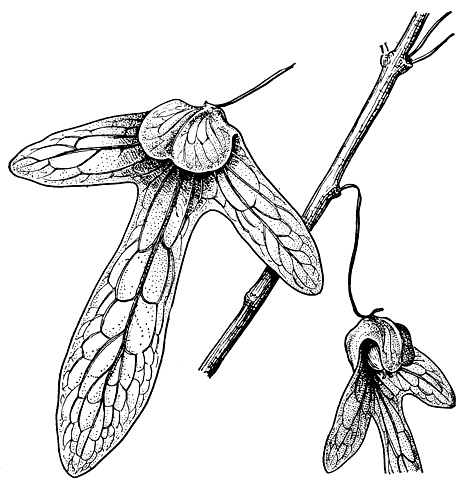
Рис. 182. Часть ветви с плодами ореомуннеи крылоплодной (Oreomunnea pterocarpa)
Возвратимся теперь к геологической летописи ореховых. В 1976 г. американские палеоботаники обнаружили новый ископаемый род ореховых - параореомуннею (Paraoreomunnea), который имеет отличия и от энгельхардии и от ореомуннеи (но ближе к последней). У па-раореомуннеи, существовавшей в среднем эоцене Северной Америки (Теннесси и Кентукки), жилкование лопастей крыла типа ореомуннеи (с некоторыми отличиями), а четвертая лопасть развита сильнее, так что она совсем скрывает плод, который оказывается в своеобразной воронке. Таким образом, родовое отличие энгельхардии от ореомуннеи косвенно поддерживается наличием предковой ископаемой формы, существовавшей к тому же в небольшом удалении от мест произрастания современной ореомуннеи. Но еще более замечательно, что ископаемые плоды, похожие на плоды ореомуннеи и параореомуннеи, обнаружены в Европе.
Для названия рода альфароя (Alfaroa), описанного в 1927 г. П. Стандли, была использована фамилия ботаника, специалиста по орхидным, тогдашнего директора Национального музея Коста-Рики А. Альфаро. Альфароя включает 6 видов, распространенных в Америке от Мексики до Колумбии. Первый вид - алъфарою костариканскую (A. costaricensis) - в 1924 г. открыл сам Стандли в горах Коста-Рики. Хотя дерево было в цвету, он принял его за хорошо известную ему ореомуннею крылоплодную, точнее, за энгелъхардию крыло-плодную (Engelhardia pterocarpa), ибо Стандли считал ореомуннею не самостоятельным родом, а лишь американской секцией рода энгельхардия.
Альфароя оказалась растением удивительно полиморфным. Листья у нее могут быть супротивными, очередными или в мутовках, листочки - супротивными и очередными, пильчатыми (у молодых экземпляров) и цельнокрай-ними или лишь частично зубчатыми (у старых экземпляров). Альфароя может быть двудомной и однодомной, сережки могут быть однополыми и андрогинными. Очень часто деревья альфарои встречаются в стерильном состоянии.
Альфароя в стадии плодоношения четко отличима от ореомуннеи, ибо плоды альфарои крупные (длиной около 6,5 см и диаметром до 3 см) и бескрылые. Женский цветок альфарои, подобно цветку ореомуннеи, снабжен трехлопастным прицветником. Но после оплодотворения пути этих цветков расходятся: у ореомуннеи прицветник вырастает в большое крыло, у альфарои остается в виде чешуи. И Стандли, встретив впервые альфарою, подумал, что ее прицветник разовьется в будущем в крыло (как и происходит у ореомуннеи).
Любопытно, что и после описания рода альфароя Стандли снова ошибся, отнеся в 1940 г, к секции ореомуннея рода энгельхардия альфарою гватемальскую (A. guatemalensis). И только в 1970 г. его ошибка была исправлена ботаниками Л. Уильямсом и А. Молиной, которые, произведя ревизию центральноамериканских ореховых, установили, что в роде альфароя 6 видов, а именно, альфароя костариканская, альфароя гондурасская (A. hondurensis), альфароя Маннинга (A. manningii), альфароя мексиканская (A. mexicana), альфароя Уилъямса (A. williamsii) и альфароя гватемальская. Альфарои - одни из самых крупных деревьев в тропиках Вест-Индии. Они не образуют чистых зарослей, встречаясь в тропическом лесу в качестве эмергентов, т. е. выступают над пологом леса.
Изучая плоды альфарои, ботаники установили, что чашечка и прицветник этого растения не принимают участия в образовании оболочки плода, как у других ореховых. Это ставит альфарою в особое положение в семействе ореховых. И, конечно, первоначальное мнение о ее близком родстве с родом орех пришлось отклонить. Видимо, правы американские ботаники У. Маннинг и Д. Стоун, считающие энгельхардию, ореомуннею и альфарою ближайшими родственниками. У. Маннинг высказывается совсем решительно. Он полагает, что неплохо было бы объединить все три рода в один. Правда, такой операции он еще не произвел. Д. Стоун, напротив, признает все три рода. Интересны его доводы в пользу большего родства ореомуннеи с альфароей, а не с энгельхардией, как считают У. Маннинг и большинство других авторов. Дело в том, что вопреки резкому внешнему несходству плодов альфарои и ореомуннеи, по анатомическому строению они очень близки друг другу.
За многие годы изучения ореховых разные ботаники предложили несколько систем семейства, и все они были основаны на типе строения плодов, вернее, на признаках величины плодов и наличия или отсутствия у них крыла. Все роды с бескрылыми плодами попадали в одно подсемейство, а роды с крылатыми плодами - в другое. Открытие альфарои и тщательное изучение ее родственных связей внутри семейства ореховых, а также мучительная проблема с платикарией, имеющей шишкоподобное женское соцветие (и соплодие) и не костянко-видный, а ореховидный плод (исключение в семействе ореховых), привели У. Маннинга, всю жизнь занимающегося ореховыми, к построению новой системы (1975) этого семейства: 1) подсемейство собственно ореховые (Juglandoideae): а) триба собственно ореховые (Juglandeae) - орех, птерокария, б) триба энгельхардиевые (Engelhardieae) - энгельхар-дия, ореомуннея, альфароя, в) триба кариевые (Саrуеае) - кария; 2) подсемейство платикариевые (Platycaryoideae) - платикария.
Эта система учитывает прежде всего строение цветка, но также совокупность других признаков, в том числе и процесс развития плодов.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'