Порядок буковые (Fagales)
Семейство буковые (Fagaceae) (Ю. Л. Меницкий)
Семейство буковых содержит 7 - 8 родов и более 900 видов, распространенных в умеренных, субтропических и тропических областях обоих полушарий, за исключением большей части Южной Америки и тропической и Южной Африки. Большинство видов семейства - листопадные или вечнозеленые деревья, часто значительной высоты, и лишь немногие - кустарники и даже кустарнички, не превышающие 30 - 40 см. Листья очередные или очень редко мутовчатые, перистонервные, с линейными, обычно рано опадающими прилистниками. Членики сосудов с лестничной, с лестничной и простой или чаще только с простой перфорацией. Цветки мелкие, однодомные, в верхоцветных соцветиях различного типа - колосовидных, сережковидных, кистевидных или шаровидных, реже одиночные. Отдельные женские дихазии у основания окружены более или менее чашевидной плюской, которая у некоторых видов нотофагуса (Nothofagus) бывает сильно редуцирована или даже полностью подавлена. Плюска образована видоизмененными конечными стерильными ветвями соцветия. Она обычно снабжена придатками разнообразной формы - чешуйками, шипами, бугорками, щетинками, гомологичными прицветникам. Число лопастей плюски обычно зависит от числа цветков в дихазии. В одноцветковых дихазиях бука (Fagus) и нотофагуса она двулопастная (2 стерильные оси 2-го порядка), в трехцветковых дихазиях кастанопсиса (Castanopsis) и нотофагуса - четырехлопастная (4 оси соцветия 3-го порядка), в семицветковых дихазиях тригоноб алану са (Trigonobalanus) - восьмилопастная (8 стерильных осей 4-го порядка), но у отдельных представителей некоторых родов число долей плюски может и не соответствовать числу цветков дихазиального соцветия.
Цветки буковых часто бывают с рудиментами органов другого пола. Чашелистиков обычно 6, реже меньше (до 2) или больше (до 8), чашевидных, более или менее сросшихся. Тычинок большей частью 6 - 12, с тонкими, свободными нитями, превышающими чашечку; пыльники интрорзные, прикрепленные к тычиночной нити у середины или у основания. Пыльцевые зерна трехбороздные или трехбороздно-поровые, у нотофагуса с 4 - 7 экваториальными бороздовидными апертурами. Гинецей из 3(2 - 9) плодолистиков; завязь 3(2 - 9) - гнездная с двумя висячими семязачатками в каждом гнезде, из которых развивается только один. Столбики свободные или сросшиеся в колонку, иногда редуцированы до коронки.
Плод - односемянный орех с твердым околоплодником, заключенный полностью или частично в деревенеющую плюску.
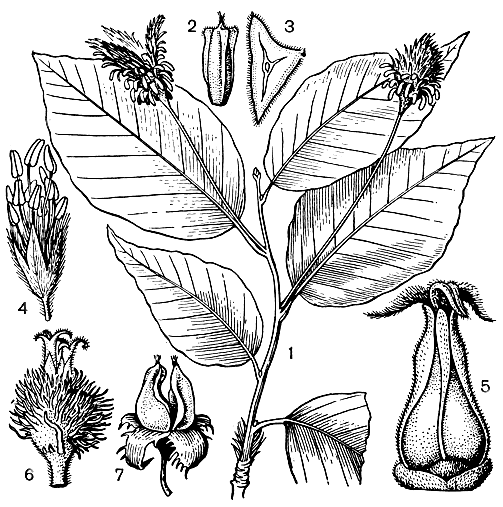
Рис. 153. Буковые. Бук Энглера (Fagus engleriana): 1 - побег с плодами; 2 - 3 - плоды (вид сбоку и в разрезе). Бук европейский (F. sylvatica): 4 - мужской цветок; 5 - женский цветок; 6 - женское соцветие; 7 - зрелые плоды
Семейство подразделяется на 2 подсемейства: буковые (Fagoideae) и каштановые (Castaneoideae). У буковых мужские цветки расположены на облиственных побегах в 1 - 3-цветковых дихазиях (нотофагус) или в дихазиальных стебельчатых головках в пазухах листьев (бук), имеют большие колокольчатые чашечки и крупные пыльники. Плюска раскрывается 2 - 4 створками, содержит 1 - 3 трехмерных (трехгранных) или двухмерных (уплощенных) ореха. Членики сосудов преимущественно или частично с лестничной перфорацией. Кроме того, для родов этого подсемейства характерен надземный тип прорастания семян (семядоли поднимаются над землей, раскрываются, зеленеют и фотосинтезируют), запасным веществом семядолей является масло. У каштановых цветки располагаются в дихазиальных пучках либо поодиночке на специализированных цветоносных побегах, или в поникающих однополых сережках, с мелкими чашечками и пыльниками. Плюска раскрывается 2 - 4 створками и содержит 1 - 3 трехгранных ореха, иногда нераскрывающаяся, с одним орехом (литокарпус - Lithocarpus); у дуба и литокарпуса от блюдцевидной до полушаровидной формы и лишь частично заключает в себе плод (желудь). Членики сосудов обычно с простой перфорацией. Прорастание семян подземное, толстые, мясистые семядоли остаются на земле и обычно служат только как источник запасных веществ (крахмала).
Бук (Fagus) - важнейшая лесообразующая порода умеренных областей северного полушария. В этом роде насчитывается 9 - 10 видов, распространенных в восточных штатах США, в горах Северной Мексики, в Европе, Малой Азии, на Кавказе и в Северном Иране, затем, после перерыва вновь появляются в континентальном Китае, на острове Тайвань и в Японии. Буки - большие листопадные деревья с колоннообразным стволом, высотой 25 - 40 м и диаметром до 1,5 м и более, покрытым серой гладкой корой, густой, округлой тенистой кроной и близко расположенными к поверхности почвы корнями; под сенью этих деревьев в сомкнутых чистых древостоях обычно имеется лишь мертвый покров. Корневая система бука без ясно выраженного стержневого корня, с боковыми корнями, имеющими уходящие вертикально вниз якорные ответвления. Мелкие окончания корней нередко направлены вертикально вверх и щеткообразно ветвятся в слое мертвой подстилки, имея хорошо развитую микоризу. У старых деревьев часто сильно разрастаются корневые лапы, в связи с чем в комлевой части ствола появляется характерная желобчатость.
Почки бука очень характерны - веретено-видные, длиннозаостренные, длиной до 1,5 см. Листья продолговато-эллиптические, с 7 - 15 парами жилок, зубчатые.
Цветки бука (рис. 153) расположены в пазухах нижних листьев и появляются одновременно с их распусканием. Мужские цветки в висячих головчатых дихазиальных соцветиях, с колокольчатой чашечкой и неопределенным числом тычинок (10 - 20) на длинных нитях. Пыльцевые зерна очень крупные, с камерными порами и очень узкими бороздами. Тяжелая пыльца бука распространяется в горизонтальном направлении на небольшое расстояние. Женские цветки в двухцветковом (в результате редукции центрального цветка) дихазиальном соцветии, с 6-лопастным околоцветником. Плюска с 4 или 3 (в случае редукции двух цветков) створками, к моменту созревания плодов сильно разрастается и древеснеет. Плод - трехгранный орех с опушенным эндокарпием, острыми (почти до короткокрылатых) ребрами. В плюске обычно по 2, реже по 4 ореха. Зародыш со складчатыми семядолями, содержащими в качестве запасного вещества до 50% масла.
Зачатки мужских соцветий бука закладываются летом предшествующего цветению года в формирующихся почках, зачатки женских - значительно позднее. У бука восточного (F. orientalis) мужские соцветия закладываются в июне, после окончания формирования кроющих чешуи; процесс формирования мужских соцветий продолжается 2 - 3 месяца, и уже в августе при вскрытии почек можно найти соцветия с зачатками пыльников. Осенью и зимой происходит дальнейшая дифференциация тканей, заканчивающаяся лишь весной. Итак, цикл развития мужских соцветий занимает 9 - 10 месяцев. В одной почке может заложиться до 6 соцветий. Зачатки женских соцветий появляются на месяц позже мужских, когда в почке уже сформированы 2 - 3 эмбриональных листа. Женские соцветия закладываются в виде меристематических бугорков в их пазухах. Большее количество зачатков закладывается при продолжительной теплой и сухой погоде, обусловливающей на следующий год большой приток углеводов к почкам и обильное плодоношение. Для нормального развития женских соцветий бук, так же как и дуб, нуждается в низких температурах зимой (минимум несколько сотен часов). Дифференциация соцветий завершается лишь весной, после установления устойчивых положительных температур. Распускание листьев и цветение бука восточного происходит в начале мая. Большинство побегов с женскими соцветиями находится на конечных частях ветвей; побеги с мужскими соцветиями располагаются по всей длине ветви, преимущественно в нижней ее части. Мужские цветки обычно раскрываются на 3 - 4 суток раньше женских. В дождливую погоду пыльники раскрываются в нижней части кроны, в солнечную - в верхней. Пыльца бука гибнет как от чрезмерной сухости воздуха, так и от избыточного увлажнения.
При созревании женских цветков рыльца сильно выдвигаются из них и выделяют липкий секрет. Для рылец даже кратковременные заморозки оказываются губительными, высокие температуры также оказывают неблагоприятное воздействие, дождь смывает липкий секрет, ухудшая как прилипаемость, так и прорастание пыльцы.
Семязачаток и зародышевый мешок заканчивают развитие только после опыления (осуществляемого главным образом ветром), так что оплодотворение яйцеклетки происходит через 2 - 3 недели. Оптимальная температура для прорастания пыльцевой трубки +18... + 22 °С. Орехи созревают в октябре. В равнинных лесах Западного Закавказья бук плодоносит почти ежегодно, на высотах свыше 1000 м бук восточный, как и бук европейский, дает обильные урожаи через 3 - 5 лет. Семена зимуют в подстилке под снегом и прорастают в марте - апреле, когда среднесуточная температура поднимается до +8... +10 °С. Семядоли, похожие на полукруглые раскрытые зеленые веера с ушками при основании, выходят на поверхность и остаются на молодом растеньице около 10 суток. С появлением первых листьев проростки становятся более устойчивыми и развивают стержневой корень длиной 20 - 30 см. В первый период развития сеянцы бука, как и дуба, нуждаются в затенении, которое в естественных условиях обычно имеется (полог деревьев), а при лесных посадках семена высевают под полог кустарников (лещина, бузина, спирея), которые впоследствии под пологом бука отмирают.
Бук живет до 500 лет, хорошо плодоносить начинает с 30 - 50 лет, дает прирост до 350 лет; наиболее быстрый рост в высоту происходит до 80 лет, после чего ствол в основном только утолщается и развивается мощная крона.
Кроме семенного размножения, в молодом возрасте бук возобновляется пневой порослью, которую дают все пни диаметром до 10 см, но уже при диаметре ствола 20 см порослевое возобновление резко снижается. Поросль из спящих почек также способна образовать стволы, особенно после рубки в хорошо освещенных местах. Корневые отпрыски редки, появляются после повреждения корней и для размножения бука существенного значения не имеют.
Бук европейский (F. sylvatica, рис. 153) является характерным элементом зоны широколиственных лесов Европы, представляет собой дерево высотой до 40 - 50 м, листья эллиптические, широко заостренные к основанию и верхушке, волнистые или слабозубчатые по краю, с 5 - 8 парами боковых жилок и шиловидными придатками плюски. Распространен почти по всей Западной Европе: его южная граница проходит через юг Болгарии и Центральную Грецию, идет вдоль Адриатического побережья Албании, Югославии, Италии, доходит до северо-востока Сицилии, проходит между островами Корсика и Сардиния и идет до Северной Испании. Северо-западная граница пересекает Великобританию, проходит по Бельгии, Нидерландам, ФРГ и Дании, не выходя, однако, к берегам Северного моря, и достигает Скандинавии. Восточная граница проходит от Калининграда через Западную Польшу, Юго-Западную Украину, Восточную Румынию, Крым и Болгарию; граница островных местонахождений бука значительно восточнее. Северная и восточная границы распространения бука определяются тепловым режимом - бук не выдерживает сокращения вегетации менее чем до 5 месяцев и снижения средней температуры января ниже -6°С.
Бук европейский - светолюбивая лесная порода и наилучшим ростом и развитием обладает при хорошем освещении, вместе с этим является одним из наиболее теневыносливых деревьев. Он устойчив к кратковременным понижениям температуры (до -35 °С), однако длительные морозы -13...-20 °С вызывают у него сильные повреждения - трещины стволов, отхождение коры. От морозов особенно страдают проростки. Бук не может жить в районах с годовым количеством осадков ниже 500 мм и низкой влажностью воздуха, предпочитает свежие и мощные почвы - на склонах с мелкими почвами имеют место массовые ветровалы. На севере ареала бук европейский растет преимущественно на равнинах, в южных районах занимает определенную высотную зону в горах - в Скандинавии он растет до высоты 190 м, в Пеннинских Альпах - до 500 м, Гарце - до 970 м, Вогезах - 1380 м, Татрах, Карпатах и Центральных Альпах - до 1400 - 1440 м, в Тироле - до 1680 м, в Пиренеях - до 1870 м, в Апеннинах - до 1970 м над уровнем моря. Нижняя граница бука может снижаться даже в южных районах его ареала в благоприятных условиях до 50-100 м над уровнем моря, что часто имеет место в Болгарии, Югославии, а также на окраине Среднедунайской равнины. В Крыму бук европейский распространен на высоте от 500 до 1300 м над уровнем моря, а отдельные его экземпляры выходят на Яйлу.
В чистых буковых лесах из-за создаваемой буком густой тени подлесок обычно отсутствует или представлен лишь тиссом и падубом; травяной покров также весьма беден - весной в буковых лесах появляются редкие эфемеры. Часто бук образует смешанные насаждения - растет вместе с грабом, дубом, черешней, кленом, явором и липой; в верхней горной зоне встречается вместе с пихтой и елью.
Бук восточный (F. orientalis) - величественное дерево, высотой до 50 м, с мощной шатро-видной кроной, продолговато-эллиптическими заостренными к обоим концам листьями с 7 - 15 парами боковых жилок; от бука европейского отличается продолговатыми листьями и более широкими нижними придатками плюски. Бук восточный распространен на Кавказе, в Малой Азии, Северном Иране и Восточной Болгарии. Ввиду того что этот вид связан переходными формами с буком европейским, его западную границу установить весьма трудно. Некоторые исследователи считают, что он распространен в Крыму и идет на запад до Греции в верхних поясах гор, в нижних поясах которых растет бук европейский. На Кавказе бук занимает почти половину всей площади, покрытой лесами. Он широко распространен на северных склонах Кавказа; в Закавказье характеризуется почти сплошным распространением, и только в верховьях отдельных рек уступает место хвойным. По Главному хребту идет от Черноморского побережья до восточной границы лесов (Шемаха), по Малому Кавказу идет на восток до реки Тертер, а на востоке снова обнаруживается в Талыше, уходя по предгорьям Эльбурса в Иран.
На Западном Кавказе бук растет почти от уровня моря, однако в других районах его кавказского ареала пояс собственно буковых лесов, где он образует чистые насаждения, простирается на высоте от 700 до 1200 м. Выше 1200 м к буку в значительной степени начинают примешиваться пихта и кавказская ель, однако верхняя граница его распространения - 2200 - 2300 м, где он входит в состав субальпийского криволесья в виде крупных кустов часто со стелющимися стволами.
Бук восточный более холодостоек, чем бук европейский. Он требователен к влажности воздуха и не может жить в районах, где относительная влажность падает ниже 70%. Лучшие бонитеты восточного бука приурочены к спокойному рельефу и глубоким почвам на высоте около 900 - 1000 м. Бук восточный обычно не заселяет мокрые почвы близ выхода грунтовых вод, недренированные впадины, размытые русла ручьев, на нижних террасах растет только при хорошем дренаже и недоступности для речных вод. Бук вынослив и к ограниченному запасу воды - может расти вместе с дубом на крутых южных склонах с мелкой почвой, однако здесь бук очень ветровален из-за поверхностной корневой системы. Возобновляется группами в просветленных местах леса - "в окнах" - при гибели деревьев; там быстро развиваются травяной покров и подлесок, необходимые буку на ранних стадиях развития, которые впоследствии гибнут при восстановлении полога букового леса.
Обычно в чистых буковых лесах подлесок отсутствует (мертвопокровные буковые леса), реже имеется редкий подлесок из боярышника, мушмулы, на Западном Кавказе - из рододендрона понтийского, рододендрона желтого, азалии, самшита, падуба, кавказской черники и папоротника орляка; травяной покров также весьма беден.
Бук американский, или крупнолистный (F. grandifolia), с хорошо выраженными изогнутыми зубцами на листьях, распространен в приат-лантических районах Северной Америки и, с 550-мильным разрывом, в горах Центральной Мексики (var. mexicana), т. е. от зоны тайги на севере и почти до субтропических лесов на юге, на равнинах или чаще на возвышенностях по берегам рек. Он образует чистые леса или растет в смеси с березой, сахарным кленом, на юге - с липой, лириодендроном и другими древесными породами.
Из двух буков, распространенных в Японии, бук городчатый (F.crenata) весьма близок к буку восточному, отличаясь от последнего яйцевидными, сверху коротко заостренными листьями. Он встречается в верхней части лесной зоны до высоты 1600 м над уровнем моря (холодноумеренная зона) к югу от полуострова Осима острова Хоккайдо, где растет с пихтой, дубами, вязами, липой, кленом и калопанаксом; травяной покров в таких лесах часто заменяет мелкий бамбук саза, иногда служащий препятствием возобновлению бука. Другой бук, распространенный в Японии, - бук японский (F. japonica) - характеризуется длинными, выступающими из плюски орехами и короткими раздвоенными придатками плюски, сидящей на длинной тонкой голой ножке. Встречается в теплоумеренной зоне Японских островов до высоты 600 м над уровнем моря в сообществе с вечнозелеными дубами, кастанопсисом и другими широколиственными деревьями.
Остальные виды бука растут в горах Центрального и Южного Китая в умеренных зонах на высотах от 1000 до 2200 м над уровнем моря, а один вид - бук Хаяты (F. hayatae) - эндемичен для острова Тайвань.
Древесина бука, приобретающая после хранения (лежания) розовато-коричневый цвет и красивый рисунок, находит широкое применение. Ее используют для изготовления мебели, бочек, колес, в строительстве, внутренней отделке помещений (паркет, облицовка), в судостроении, для подводных сооружений. Из нее также изготовляют фанеру, музыкальные инструменты, ружейные ложа; годится она и для декоративного оформления радиоприемников и телевизоров. Древесину бука используют для получения древесного угля, уксуса и метилового спирта, ценится также и буковый деготь. Однако она малопригодна для наружных работ - на открытом воздухе легко разрушается грибами; в жаркое время года сырая древесина быстро портится, буреет, по ней распространяется мраморная гниль. Для устранения этих недостатков широко используется пропитка древесины различными химикатами. Возможна пропитка деревьев на корню органическими и неорганическими красителями для получения красивой текстуры древесины. При этом приобретается устойчивость к гниению и увеличивается предел прочности древесины.
Орехи, урожай которых может достигать 450 кг/га и до 8 кг с дерева, содержат 30 - 50% жира и до 30% белка; это излюбленный корм для кабанов и свиней. Из орехов получают светло-желтое, приятное на вкус масло, мало уступающее прованскому и используемое в пищевой промышленности и технике. Сырые орехи ядовиты из-за содержащегося в них алкалоида фагина, вызывающего головные боли у людей и выкидыши у коров. В вареном виде орехи используют в корм птице, а также крупному и мелкому рогатому скоту, но не пригодны в качестве корма для лошадей. Побеги бука представляют хороший корм для скота, годятся они и для силосования.
Виды нотофагуса (Nothofagus, рис. 154) - большие деревья, в благоприятных условиях влажности и температуры достигающие в высоту 40 - 50 м и в диаметре 1,5 - 2 м, часто с угловатым стволом, иногда имеющим контрфорсы у основания. Нотофагусы, растущие в горах, - небольшие деревья, образующие редколесья и кустарниковые заросли. Нотофагусы являются основными лесообразующими породами антарктической части Южной Америки (к югу от 33° южной широты) и прилегающих к ней южно-тихоокеанских субантарктических островов, Западной Австралии, Тасмании, Новой Зеландии, Новой Каледонии и Новой Гвинеи (включая острова Д'Антркасто, Гуденаф, Норманби и Новая Британия), где растут почти от уровня моря (Новая Каледония) до границы вечных снегов в горах (карта 15). Всего насчитывается около 40 видов нотофагусов, из них половина растет в Новой Гвинее и Новой Каледонии, а остальные - в Южной Америке, Австралии и Новой Зеландии.
Побеги нотофагусов обычно округлые, у многих видов зигзагообразные, голые. Почки с крестообразно налегающими чешуями, в отличие от почек бука, короткие, притуплённые, яйцевидные, с чешуями, расположенными в 4 четкие ортостихи.
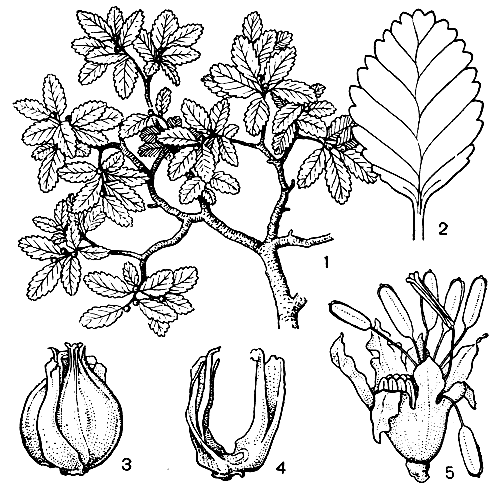
Рис. 154. Нотофагус карликовый (Nothofagus pumilio): 1 - побег с плодами; 2 - лист; 3 - плоды с плюской; 4 - плюска; 5 - мужской цветок
У 6 листопадных видов секции нотофагус (Nothofagus), к которой относятся 7 - 8 южноамериканских видов и 1 тасманийский, листья мягкие, в почках согнутые вдоль боковых жилок, городчатые или зубчатые по краю. У вечнозеленых видов (секция калюспарассус - Calusparassus) Австралии, Новой Зеландии и умеренной зоны Южной Америки листья кожистые, голые или в различной степени опушенные, цельнокрайние или мелкозубчатые по краю, усеяны смоляными железками. Новогвинейские вечнозеленые виды, выделяемые иногда в особый род трисингине (Trisyngyne) вместе с новокаледонскими видами, отличаются от других нотофагусов крупными щитковидными прилистниками, усеянными, как и почечные чешуи, смоляными железками, и располагающимися на побеге в 4 хорошо выраженных ряда. Листья вечнозеленых видов согнуты в почках вдоль средней жилки верхней поверхностью внутрь; молодые листья новогвинейских видов покрыты толстой смолистой пленкой. На листьях листопадных видов иногда также присутствуют смоляные многоклеточные дермальные желёзки на одной или обеих сторонах.
Соцветия нотофагусов - 1 - 3(7) - цветковые дихазии, расположенные в пазухах листьев, причем мужские соцветия находятся обычно у основания, а женские - на вершине побега; иногда мужские и женские соцветия располагаются раздельно на различных побегах одного дерева. Мужские цветки на коротких изогнутых цветоножках, с колокольчатой или воронкообразной чашечкой, обычно с многолопастным или усеченным отгибом и 8 - 40 тычинками с линейно-продолговатыми остроконечными сверху, крупными (длиной 1 - 8 мм) пыльниками, прикрепленными к нитям своими основаниями. У нотофагусов чрезвычайно своеобразная пыльца с 4 - 7 экваториальными бороздо-видными апертурами, отличающаяся от пыльцевых зерен других буковых.
Женские цветки в 1 - 3 (редко 7) - цветковых дихазиях на коротких ножках, в пазухах верхних листьев побега, окруженные плюской. Характерной особенностью нотофагусов, в отличие от буков, является присутствие в подавляющем большинстве случаев центрального (всегда двухмерного) цветка дихазия, ввиду чего прицветные листья первого порядка всегда развиты. Плюски нотофагусов очень разнообразны: 2-, 3- и чаще 4-створчатые, реже они редуцированы до коротких малозаметных придатков, скрытых в прицветниках, или отсутствуют. Обычно плюска снаружи покрыта кольцеобразно расположенными чешуевидными придатками - пластинками. Для плюсок антарктических видов особенно характерны стебельчатые железки, представляющие собой большие (длиной 0,5 - 2 мм) многоклеточные структуры на концах перистораздельных пластинчатых придатков. Стеблевидная часть железки имеет морфологические черты модифицированного листового сегмента; головчатая железистая часть органа состоит из слоя столбчатых секреторных клеток эпидермального происхождения, окружающих губчатую сердцевину.
Чашечка женских цветков из 6 (у трехмерных), 4 (у двухмерных) мелких кожистых листочков, расположенных в 2 круга, иногда редуцирована.
В отличие от семязачатков всех остальных родов семейства виды нотофагуса имеют простой интегумент. Плод - обычно крылатый орех, сходный по внешней морфологии с орехом бука; эндокарпий, в отличие от последнего, голый.
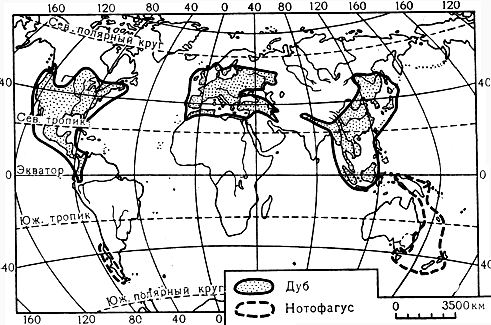
Карта 15. Ареалы родов дуб и нотофагус
Период цветения у нотофагусов Старого Света совпадает с влажным сезоном (октябрь - март), хотя сильные дожди не благоприятствуют опылению. Опыление продолжается 1 - 2 недели, после чего мужские цветки опадают и дерево создает иллюзию женского экземпляра, в результате чего некоторые виды долгое время считали двудомными. Опыление может происходить при переносе ветром не только пыльцы, но и цветков или даже целых дихазиев. Продукция пыльцы иногда столь значительна, что, например, в Новой Зеландии над нотофагусовым лесом в период цветения висит и передвигается облако, как в Европе над ржаным полем; ветреный климат океанических островов способствует дальнему ее разносу и образованию гибридов.
У нотофагусов, как и у бука, существуют ярко выраженные урожайные и неурожайные годы. Семена созревают приблизительно через полгода после опыления. Они меньше семян буков и только у немногих видов не уступают им по размерам и, по-видимому, съедобны. Новогвинейские виды производят мало плодов, и многие из них нежизнеспособны.
Сеянцы нотофагусов отличаются от сеянцев буков еще и присутствием на гипокотиле и краях семядолей железистых головчатых волосков.
Плоды новогвинейских и новокаледонских видов не выдерживают контакта с морской водой и не могут поэтому переноситься ею (кроме того, они тонут в воде); распространение плодов происходит орехоядными птицами, ветром, ручьями на небольшое расстояние; вообще семенное возобновление нотофагусовых лесов в этом районе происходит очень медленно. После пожаров обычно не появляется прикорневой поросли, однако поваленные деревья в сырых мшистых лесах дают вертикальные укореняющиеся побеги. Эктомикориза играет существенную роль в минеральном питании нотофагусов; возможно, благодаря ей они способны поддерживать свое доминирование в лесах.
По склонам Анд (Чили, Патагония) и прилегающим к Чили островам нотофагус распространен в полосе от 33 до 56° южной широты и до восточного побережья Огненной Земли в умеренно теплой и холодной зонах с годовым количеством осадков 500 - 2000 мм, среднегодовыми температурами + 5... + 13 °С и слабым различием летних и зимних температур; виды этого рода являются весьма обычными эдификаторами холодных, вечно сырых мшистых лесов, которые ввиду зимнезелености многих видов имеют одинаковый облик во все времена года.
Самый северный вид - нотофагус косой (N. obliqua) - ранее являлся одним из наиболее распространенных деревьев в девственных лесах, прежде покрывавших склоны Анд на широте около 35° южной широты. Это высокое листопадное дерево с дважды зубчатыми листьями, встречающееся в Чили к югу от реки Мауле обычно в сообществе с нотофагусом Домбе (N. dombei), причем эти 2 вида дают основной строевой лес Чили. В горных лесах в окрестностях вулкана Чильян (37° южной широты) нотофагус косой поднимается до высоты 1500 м, а на больших высотах сменяется также листопадными нотофагусом антарктическим (N. antarctica) и нотофагусом карликовым (N. pumilio, рис. 154). Нотофагус антарктический, по-видимому, самый широко распространенный высокогорный вид, встречающийся как на чилийской, так и на аргентинской стороне Анд от 38° южной широты до Магелланова пролива и Огненной Земли. На Огненной Земле он доминирует в восточной, более сухой части острова; в других местах растет в верхней лесной зоне, обычно в сообществе с араукарией на высоте до 1100 м над уровнем моря. Замечательно, что ареалы родов нотофагус и араукария в значительной мере совпадают как в Старом, так и в Новом Свете. В альпийской зоне - у вечных снегов - нотофагус антарктический представлен карликовой формой и растет там вместе с барбарисом, дримисом, пильгеродендроном.
Листопадный вид нотофагус карликовый, имеющий сильно редуцированные женские дихазии, мелкие, почти округлые, тупые листья с двойными зубцами и распростертый ствол с восходящими ветвями и бородавчатыми веточками, распространен в районе Магелланова пролива в горах на границе вечных снегов.
К югу от широты острова Чилоэ нотофагус замещает многие виды деревьев, хотя на крайнем юге Огненной Земли и на более южных островах он редко поднимается на значительные высоты. В районах, прилегающих к Магелланову проливу, почти единственным источником топлива служит древесина нотофагуса березовидного (N. betuloid.es). В благоприятных условиях это большое дерево со стволом в окружности до 6,5 м; нотофагус березовидный обладает мелкими (длиной до 1,5 см), вечнозелеными, эллиптическими, тупыми, городчато-зубчатыми, толстыми листьями, снизу покрытыми точечными смоляными желёзками, напоминающими листья антарктической березы. На Огненной Земле это главная лесообразующая порода, однако там нотофагус, березовидный растет корявым деревом с расставленными сильно искривленными сучьями.
Нотофагус образует большие леса во многих районах Южного острова и южной части Северного острова Новой Зеландии. Он встречается на различных высотах - почти от уровня моря и до вершин гор как во влажных, так и в сухих местах, образуя густые леса и заросли, почти без подлеска. В горах Северного острова до высоты 1150 м распространен вечнозеленый нотофагус бурый (N, fusca) - высокое дерево (до 30 м) со стволом, достигающим в окружности 10 м; листья у него кожистые, колючезубчатые, имеющие по 5 - 6 пар жилок; на вершинах гор и в других неблагоприятных условиях растет в виде мелкого кустарника. Этот вид является аналогом нотофагуса березовидного Огненной Земли.
В горных районах Австралии и острова Тасмания распространены нотофагус Каннингема (N. cunninghamii) и нотофагус Гунна (N.gunnii). Первый из них - густо облиственное склерофилльное дерево с вечнозелеными треугольными мелкими листьями, снабженными 3 - 4 парами боковых жилок, идущих в зубцы. В верхнем горном поясе острова Тасмания и в горах Юго-Восточной Австралии этот вид образует густые, обычно невысокие, кустарниковые заросли. Второй вид в горах Тасмании образует непроходимые заросли на высотах 1450 - 1625 м над уровнем моря, ввиду того что его ветви обычно изогнуты под давлением массы выпадающего там снега. Севернее 28° южной широты в Австралии (Квинсленд и Новый Южный Уэльс) встречается нотофагус Мура (N. moorei). Это вечнозеленое дерево, стволы которого обычно густо покрыты эпифитами, доминирует в местных лесах, успешно конкурируя с эвкалиптами.
На островах Новая Каледония и Новая Гвинея (вместе с прилегающими островами Д'Антркасто и Новая Британия) растет около 20 видов вечнозеленых нотофагусов с двухраздельной плюской (подсекция Bipartitae). Виды этой подсекции иногда выделяют в особый род трисингине (Trisyngyne).
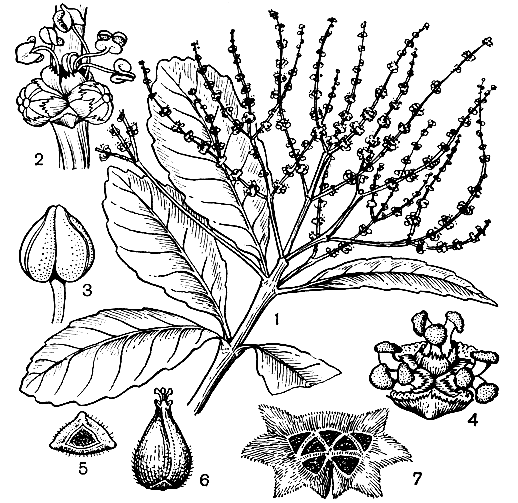
Рис. 155. Тригонобаланус мутовчатый (Trigonobalanus verticillata): 1 - побег с мужскими соцветиями; 2 - мужской дихазий; 3 - тычинка; 4 - женский дихазий; 5 - 6 - плод (вид снизу и сбоку); 7 - плюска с рубцами от 7 орехов
В горных дождевых лесах Новой Гвинеи виды нотофагуса встречаются на высотах от 1000 до 3100 м над уровнем моря, немногие из них растут и в более низких поясах гор. На высотах 1500 - 3000 м виды нотофагуса являются доминантами буколаврового мшистого леса, включающего в качестве важнейших компонентов также кастанопсис, литокарпус, араукарию, либоцедрус, эвгению, подокарп, энгельхардию и др. Пять новокаледонских видов этой секции в противоположность высокогорным новогвинейским часто растут в низменных участках острова на высотах 150 - 250 м над уровнем моря, хотя распространены и в горных лесах до высоты 1350 м. На верхней границе распространения новогвинейские и новокаледонские виды, подобно новозеландским и южноамериканским родственникам, часто становятся карликовыми деревьями и даже кустарниками.
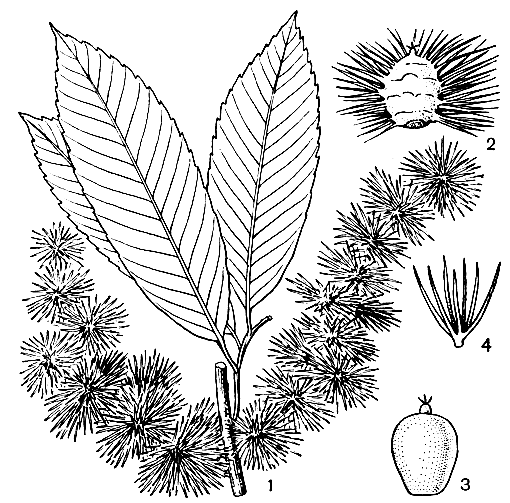
Рис. 156. Кастанопсис хобойский (Castanopsis choboensis): 1 - часть побега с плодами; 2 - плюска; 3 - орех; 4 - шип плюски
Древесина нотофагуса по механическим свойствам похожа на древесину бука. Поэтому в странах южного полушария она находит аналогичное применение и используется в строительстве, в экипажном производстве, для изготовления мебели, фанеры, для получения древесного угля, после импрегнации годится для телеграфных столбов железнодо рожных шпал и т. д. Древесина некоторых видов благодаря красивой текстуре и цвету используется краснодеревцами. Древесина нотофагуса косого устойчива к воздействию влажного климата и без пропитки может применяться для строительства мостов и изготовления шпал. В Новой Гвинее папуасы используют некоторые виды в качестве орнаментального дерева, высаживая их на фермах, вдоль дорог и садов. Виды нотофагусов успешно культивируются в Европе, главным образом в Великобритании. Замечательный род тригонобаланус (Trigonobalanus, рис. 155), впервые обнаруженный сравнительно недавно в джунглях горы Кинабалу на северо-востоке острова Калимантан, обладает наиболее примитивным в семействе строением женского дихазия и плюски и в полной мере, несмотря на некоторые черты специализации других структур, может быть назван живым ископаемым. В настоящее время известны только 3 вида этого рода: тригонобаланус мутовчатый (Т. verticillata, рис. 155), растущий на юге полуострова Малакка, севере острова Калимантан и на острове Сулавеси, и тригонобаланус дойчангский (Т. doichangensis), произрастающий на севере Таиланда. Недавно третий вид был открыт в тропической Америке. Крупные экземпляры тригонобалануса мутовчатого внешне по форме кроны похожи на старые подстриженные буки.
Если у тригонобалануса дойчангского имеется очередное листорасположение, то у тригонобалануса мутовчатого мы встречаемся с четко выраженным мутовчатым, неизвестным доселе ни у одного представителя семейства: волнистозубчатые листья этого вида длиной 5 - 10 см и шириной 3 - 4 см, с 6 - 8 парами боковых жилок образуют мутовку из 3 листьев, с попарно сросшимися в каждом секторе прилистниками.
Тригонобаланус мутовчатый растет в среднегорном (субтропическом) поясе полуострова Малакка, островов Калимантан и Сулавеси (дождевой горный лес) на высотах 850 - 1500 м над уровнем моря. Тригонобаланус дойчангский известен только из северных районов Таиланда (провинция Чиангмай), где является обычным деревом в среднегорной зоне на высотах 1200 - 1765 м над уровнем моря.
Почти все виды рода кастанопсис (Castanopsis, рис. 156) - вечнозеленые деревья высотой до 40 м, в неблагоприятных условиях - кустарники, распространенные, за исключением одного калифорнийского вида, в Восточной и Юго-Восточной Азии от Гималаев до Японии, от северной половины острова Хонсю до Новой Гвинеи, главным образом в тропических, субтропических и реже в теплоумеренных поясах гор; в последних двух зонах очень часто являются основными лесообразующими породами. В роде насчитывается около 120 видов.
Листья кастанопсисов кожистые, зубчатые или цельнокрайние. Верхушечные почки на побегах обычно имеются. Соцветия кастанопсисов прямостоячие, колосовидные или в метельчатых пучках, обычно однополые; мужские - более длинные, расположены в пазухах листьев ниже более коротких и толстых женских соцветий. Мужские цветки в 1 - 3-цветковых дихазиях, с мелкой 3-членной 2-круговой чашечкой со сросшимися у основания чашелистиками; тычинок 6 - 12, с длинными нитями и почти шаровидными или яйцевидными пыльниками, подвижно прикрепленными к нити спинкой, с полушаровидным пистоллодием в центре. Пыльцевые зерна сходны с пыльцой литокарпусов, мелкие, шаровидные или эллипсоидальные, без резко выраженных особенностей скульптуры. Женские цветки в 1 - 3-цветковых дихазиях, окружены 2 - 4-створчатыми плюсками разнообразной формы, с кувшинчатыми 2-рядными 3-членными чашечками, доли которых срастаются у основания, оставляя свободным 6-лопастный короткий отгиб. Часто имеется 6 стаминодиев, противостоящих долям чашечки. Плюска обычно асимметричная, молодая закрытая, полностью скрывающая дихазий, так что снаружи торчат только рыльца, при созревании раскрывающаяся или вообще незамкнутая. Она покрыта колючками, шипами, поперечными гребнями, бугорками, чешуйками. У большинства видов плюска четырехстворчатая, покрытая шипами, содержит 3, иногда 1 - 2 ореха. У более продвинутых видов 2 или 4 створки плюски полностью срастаются и не раскрываются при созревании плодов по швам. У ряда видов плюска почти полностью срастается с содержащимся в ней одиночным орехом.
Некоторые особенности представляет плюска североамериканского кастанопсиса золотисто-листного (С. chrysophylla), иногда выделяемого в особый монотипный род хризолепис (Chrysolepis). Это дерево высотой 20 - 30 м, с вечнозелеными продолговато-ланцетными листьями, покрытыми снизу рыжеватыми чешуйчатыми волосками, и обоеполыми соцветиями, обитающее на сухих песчаных и скалистых почвах в долинах и на склонах каньонов в области произрастания секвойи. Плюска этого специализированного вида (или рода), внешне (когда покрыта шипами) очень похожая на плюски других кастанопсисов, имеет свободные сегменты, что, однако, нельзя в данном случае рассматривать как примитивный признак. Орехи кастанопсисов созревают на первом или втором году после опыления.
В систематическом отношении род кастанопсис разделяется на несколько секций. Наиболее богатая видами секция кастанопсис (Castanopsis) со свободными (не сросшимися со стенками плюски) орехами и плюсками, покрытыми со всех сторон, кроме адаксиальной, иглами, как у каштана. Сюда относятся наиболее известные виды - кастанопсис индийский (С. indica), кастанопсис серебристый (С. argentata), кастанопсис Делавэ (С. delavayi) и многие другие.
Кастанопсис индийский - дерево высотой до 30 м и диаметром ствола до 90 см, с крупными (длиной до 24 см), кожистыми, обратнояйце-видными, острозубчатыми листьями; плюски округлые, диаметром 2 - 4 см, с длинными густыми иглами, занимающими почти всю их поверхность. Кастанопсис индийский растет в субтропических и теплоумеренных районах Гималаев (от Непала до Бирмы), в горах Южного Китая (от Юньнаня до острова Хайнань), на острове Тайвань, в Таиланде и Вьетнаме, образуя густые заросли на красных и глинистых почвах. Быстро растущее дерево с красноватой или коричневой древесиной, прочной, плотной и гибкой, которая широко используется для строительства и всевозможных поделок, хорошо сохраняется в воде и поэтому применяется для сооружения свайных построек. Древесина почти не разрушается термитами. Кора содержит танины. Плоды, урожай которых почти всегда очень обилен, вполне съедобны.
На плодородных вулканических почвах островов Ява, Калимантан, Суматра, на полуострове Малакка, в Бирме и Ассаме часто встречается кастанопсис серебристый - дерево высотой до 25 м, с серебристыми снизу, тонкокожистыми, эллиптическими, оттянутыми к верхушке и к основанию листьями и шаровидными шиповатыми плюсками диаметром около 3 см, содержащими по 3 ореха. На Яве это дерево цветет в августе - октябре и плодоносит в декабре - январе. Древесина широко используется особенно в густонаселенных районах на западе Явы для построек, плоды съедобны.
Хорошую строительную древесину и съедобные плоды дает кастанопсис Делавэ, растущий в горных лесах Западного Китая не выше 2400 м большими зарослями, в благоприятных условиях достигает в высоту 25 м, но часто встречается в виде кустарника с голой черной или пепельно-серой корой и жесткими, тупыми, мелкозубчатыми, широкоэллиптическими листьями, беловато-серебристыми снизу. Имеет асимметричную плюску диаметром около 2 см, с одним орехом, обычно с 4 концентрическими линиями и редко расставленными по ней острыми шиловидными колючками, срастающимися у основания.
Виды секции псевдопазания (Pseudopasania) характеризуются тонкостенными плюсками без длинных шипов, покрытыми волнистыми рядами чешуек или бугорков, и обычно с одним свободным орехом.
В Восточных Гималаях, в горах Ассама, Бирмы и Лаоса широко распространен кастанопсис ланцетолистный (С. lancifolia) - маленькое дерево с цельнокрайними ланцетными, заостренными у верхушки, слегка рыжеватыми от опушения снизу листьями и очень твердой древесиной. Плюска полностью закрывает орех, при созревании которого разрывается на верхушке неправильными долями.
Самый северный вид рода - кастанопсис длинноостр оконечный (С. cuspidata) - встречается как в диком виде, так и в культуре в Японии (острова Хонсю и Кюсю), Корее, Китае (провинции Цзянси, Цзянсу, Гуандун), на острове Тайвань. Это невысокое (10 - 25 м) дерево с гладкой коричневой корой и кожистыми яйцевидно-ланцетными заостренными листьями. Плюска этого вида слегка асимметрична, при созревании плодов открывается 2 - 4 створками. Плоды съедобны; их едят жареными во время религиозных праздников, а также используют для лечебных целей; кору применяют для окраски тканей и ниток.
У видов секции каллеокарпус (Calleocarpus) плюска очень толстая, в значительной степени, а иногда и полностью сросшаяся с перикарпием ореха, снаружи несет иглы или, чаще, шиповатые или чешуйчатые волнистые кольца. Орех с очень большим рубцом (иногда рубец покрывает весь плод).
В тропических лесах Суматры, Калимантана, полуострова Малакка, Филиппин встречается кастанопсис гладкоплодный (С. inermis) - дерево высотой до 30 м, с очень жесткими, яйцевидными, коротко заостренными на верхушке, серебристыми или рыжеватыми снизу листьями, грушевидной 2 - 3-лопастной плюской с 3 - 5 рядами гребней, образованных короткими пирамидальными буграми. Плюска содержит от 1 до 3 плодов, причем средний - самый мелкий и трехгранный. Древесина прочная, светло-коричневая, горьковатая на вкус, с мышиным запахом.
Род каштан (Castanea) является северной ветвью секции кастанопсис рода кастанопсис и весьма близок к последнему роду; некоторые авторы и в настоящее время объединяют оба эти рода в один. Действительно, нет ни одного вполне устойчивого и существенного признака (по-видимому, кроме летнезелености многих каштанов), который был бы характерен только для этого рода, однако можно выделить ряд тех, которые преобладают у каштана и менее часто встречаются у кастанопсиса. Это, помимо вышеупомянутой летнезелености, присутствие у него обоеполых соцветий, 6 - 9-гнездной завязи, симметричность плюски.
Каштаны - большие, высотой до 35 м, листопадные деревья или кустарники, иногда это карликовые кустарники высотой до 30 см. Около 11 - 12 морфологически довольно близких и иногда трудноразличимых видов каштана распространены, как и виды бука, в Средиземноморье, на Кавказе, в Восточных Гималаях, Восточной Азии и приатлантической Северной Америке.
Верхушечная почка отсутствует. Листья эллиптические, острозубчатые, с многочисленными параллельными боковыми жилками, покрыты снизу микроскопическими головчатыми железками, с парой опадающих прилистников. Цветки в колосовидных соцветиях. Мужские цветки на жестких прямостоячих осях в 3 - 7-цветковых дихазиях, с 6 - 20 тычинками; строение пыльников и пыльцевых зерен сходно с таковым у кастанопсиса. Женские дихазии по 1 - 3 в основании обоеполых соцветий. Плюска у каштана, в отличие от кастанопсиса, появляется уже во время цветения, вполне развитая, покрытая, за исключением шовных зон, только ветвистыми шипами, при созревании плодов раскрывается 4 или 2 створками. Плоды - орехи (каштаны) яйцевидной формы, иногда деформированные благодаря сжатию в плюске, с тонким сухим околоплодником и широким плодовым рубцом при основании (рис. 157).
Время цветения европейско-кавказского каштана настоящего, или посевного (С. sativa, табл. 40), зависит от высоты местности и происходит с конца мая до начала июля; у американского каштана зубчатого - с конца июня до начала июля; у американского каштана карликового - с конца мая до конца июня. Мужские соцветия опадают сразу после отцветания, но у обоеполых соцветий мужская часть остается до полного созревания плодов. Каштаны, как и прочие роды семейства, - растения однодомные, но, как исключение, встречаются экземпляры с соцветиями только одного пола. Такие деревья могут давать плоды, если по соседству имеются экземпляры с цветками противоположного пола.
Последовательность функционирования полов каштана посевного бывает различной: одни деревья являются протандричными, другие - протогиничными, и пыльца последних служит для опыления первых, и наоборот. Цветки американского каштана зубчатого обнаруживают двойную дихогамию: нижние соцветия цветоносных побегов с мужскими цветками развиваются первыми, а женские цветки у основания более молодых соцветий, расположенных выше на цветоносном побеге, цветут в следующую очередь, а еще позже наступает цветение другой партии мужских цветков, расположенных на тех же обоеполых соцветиях. Одни авторы считают, что функцию опыления выполняют позднораспускающиеся цветки второго поколения, другие, а это более вероятно, что рано раскрывающиеся мужские цветки доставляют пыльцу для рано созревших женских, позднее - для запоздавших в своем развитии.
Иногда одиночно стоящие деревья оказываются бесплодными, требуя, по-видимому, для оплодотворения перекрестного опыления. Исключение составляет лишь низкорослый американский каштан карликовый, он может размножаться и партеногенетически, т. е. развивать нормальные плоды без всякого опыления.
Несмотря на обилие производимой каштаном пыльцы, он, по-видимому, является более энтомофильным, чем анемофильным растением, о чем свидетельствует небольшая, в виде маленькой точки, поверхность слегка клейкого рыльца и большое количество насекомых, посещающих его ароматные соцветия, - пчел, мелких мух, жесткокрылых, в основном потребляющих его пыльцу. Пчелы в каштановом лесу собирают много меда, несмотря на небольшое количество нектара, выделяемого мужскими цветками.
Период созревания плодов у каштана посевного продолжается с конца сентября до середины ноября, у каштана зубчатого - с конца августа до начала сентября, у каштана карликового - с конца сентября до конца октября. При созревании плодов орехи обычно выпадают из треснувшей плюски, но иногда опадают в плюсках или даже с осями соцветий. Прорастают орехи весной или в начале лета.
На корнях каштана, как и на корнях других буковых, имеется эктомикориза. Каштан настоящий плодоносит с 15 - 40-летнего возраста, зубчатый - с 20-летнего, а карликовый начинает давать плоды уже с 2 - 4-летнего возраста. Полноценное плодоношение каштана настоящего наступает обычно в возрасте 60 лет, причем обильные урожаи наблюдаются каждые 2 - 5 лет в зависимости от условий обитания.
Каштаны часто достигают огромных размеров и доживают до 1000 лет и более: были описаны деревья до 26 м в окружности. Знаменитый каштан Этны, состоявший из 5 стволов, имел 64 м в окружности и возраст около 3000 лет.

Рис. 157. Плоды каштана настоящего (Castanea sativa)
На Кавказе большие деревья каштана достигают в высоту 35 м и в диаметре 2 м, их возраст в среднем 200 - 300 лет; изредка в горных лесничествах Краснодарского края встречаются экземпляры диаметром до 2,5 м, возраст которых составляет 600 - 800 лет. Вполне здоровыми деревья бывают, однако, лет до 100, после чего их сердцевина начинает гнить и появляются дупла.
Каштаны сильно страдают от грибковых заболеваний, самым страшным из которых является завезенная в Америку из Китая в начале XX в. болезнь коры и ствола, вызываемая грибком Endothia parasitica. Этот грибок уничтожил почти все насаждения американского каштана зубчатого.
Для нормального развития каштан требует непрерывной теплой погоды, но не выносит сильной жары и длительной засухи. Цветение каштана происходит при наступлении устойчивой температуры +15... + 18°С, а для полного созревания плодов средняя температура сентября должна быть около +15 °С, а октября - около +9 °С.
Северной границей произрастания каштана настоящего, где он еще плодоносит, считают изотерму января -1°, которая в Европе проходит через Шербур, Руан, Реймс, Франкфуртна-Майне, Прагу, Краков, Львов, Киев. В Европе каштан посевной не выносит длительного понижения температуры ниже -15 °С, но на Кавказе он несколько более холодостоек - прекрасно развивается на Сурамском хребте,, где температура падает ниже -16 °С, а в Краснодарском крае переносит кратковременные морозы до -25 °С. Японский каштан городчатый выдерживает еще более сильные морозы, нормально развиваясь на юге острова Хоккайдо. Наиболее морозостойким является американский каштан зубчатый, выдерживающий морозы до -27 °С.
Кроме тепла, каштан требователен к влажности воздуха и почвы, лучше всего он развивается в областях, где выпадает не менее 1000 мм осадков и относительная влажность воздуха достигает 60 - 70%; большее количество осадков не приносит каштану вреда.
Обычно каштан избегает известковых почв, а также плотных глинистых, болотистых или кислых, встречаясь на почвах, образованных на гранитах, гнейсах, порфирах, сланцах и песчаниках.
Каштан представляет относительно теневыносливую породу, занимая среднее положение между дубом и буком. Обычно каштан приурочен к склонам затененных румбов и избегает сильно освещенных.
Благодаря хорошо развитой и глубоко идущей корневой системе каштан хорошо противостоит ветровалу, но избегает открытых мест из-за того, что сильно страдает от иссушающих ветров, которые особенно вредят ему во время цветения.
В большинстве местообитаний каштан растет вместе с буком, грабом, дубом, ясенем, ольхой, пихтой. При произрастании вместе с буком быстрый рост каштана в первые годы позволяет ему обогнать бук, который служит ему хорошим подгоном, но в дальнейшем бук может сменить каштан, темп роста которого сильно замедляется к 50 годам, если не будет каких-либо благоприятствующих каштану условий.
Каштан посевной - дерево высотой до 35 м, с огромной шатровидной кроной, с крупными опушенными остроконечно-зубчатыми листьями и плюской, содержащей 3 ореха, раскалывающейся четырьмя створками при их созревании. Он распространен по всей Южной Европе и Средиземноморью, растет в Западном Закавказье и по южным склонам Главного Кавказского хребта. В Средней и Северной Европе широко культивируется, но не плодоносит; самое северное место произрастания - в Норвегии (63° северной широты), где он растет в виде крупных кустов. Очень давно культивируется в Европе, где поэтому трудно точно установить его естественный ареал. В Испании и Португалии вместе с дубом и буком образует большие леса, поднимаясь в горы до высоты 1600 м над уровнем моря. В Италии каштан очень обилен: на юге этой страны и на юге острова Сицилия он поднимается в горы до высоты 1200 - 1500 м над уровнем моря. В Средней Европе каштан растет обычно на равнинах, в более южных районах постепенно поднимаясь в горы.
Древесина каштана настоящего внешне похожа на древесину дуба, но менее темная, не темнеет от контакта с воздухом, эластичная, средней твердости, прочная, с красивым рисунком, но легко колется. Ее используют для строительства домов, производства мебели, внутренней отделки помещений, в экипажном деле, в судостроении (мачты); особенно ценятся каштановые винные бочки - ввиду своей непроницаемости каштановая клепка прекрасно сохраняет вино от испарения, а также не вносит в него посторонних примесей. Молодая поросль и тонкие ветки идут на изготовление плетеных изделий, низкоствольник - на шесты для хмеля и тычины для винограда. Молодые побеги и листья являются хорошей добавкой в корм животным. Древесина содержит много танинов. Плоды каштана играют значительную роль в питании населения большинства стран Южной Европы, а в некоторых местностях (остров Корсика) фактически заменяют хлеб. Плоды каштана используют в пищу сырыми, вареными, печеными, жареными. Из сушеных каштанов приготовляют муку, по питательности почти не уступающую пшеничной; обычно ее подмешивают к пшеничной, ржаной или кукурузной муке для выпечки хлеба и лепешек. Значительный процент каштанов используется в кондитерском деле, где они являются составной частью пирожных, тортов, пирогов и начинок для конфет. Поджаренные каштаны употребляют как суррогат кофе. В тех местах, где каштаны находятся в изобилии, они служат для откорма домашней птицы и животных.
В Западной Европе существует до 500 сортов каштанов, многие из которых были завезены туда еще 2000 лет назад из Малой Азии. Сорта каштанов сильно различаются по величине плодов, их массе, содержанию Сахаров, толщине кожуры, химическому составу. Если обычные сорта каштанов содержат в плюске по 2 - 4 сравнительно небольших ореха, то марроны - улучшенные сорта - имеют по 1 - 2 очень крупных ореха хорошего вкуса. Деревья, производящие марроны, более требовательны к почве, менее урожайны и имеют древесину худшего качества, чем ординарные сорта. Орехи лучших сортов марронов достигают в диаметре 4 см.
Американский каштан зубчатый (С. dentata, рис. 158) - дерево высотой до 35 м, с голыми зубчатыми листьями, имеющими клиновидные основания. Этот вид образовывал большие леса в горах и на равнинах востока США - от штата Мэн (43° северной широты) и южных берегов озера Онтарио до Миссисипи, Луизианы, Алабамы, где он рос на высоте 400 - 1200 м над уровнем моря. До инвазии Endothia parasitica эти леса составляли около 1/10 всех лесов США. В настоящее время вид сохранился только в виде поросли в отдельных отмирающих древостоях, древесина которых сейчас используется только для производства дубителей.
Японский каштан городчатъш (С. crenata) - небольшое дерево, высотой до 15 м, с ланцетными тонкозубчатыми листьями; плюски с почти голыми колючками длиной до 3 - 5 см содержат по 3 ореха. Растет в Восточном Китае и Японии на островах Кюсю и Хонсю: на юге - в диапазоне высот 500 - 1500 м, в северных районах Хонсю - на высоте 200 м над уровнем моря. Наилучший рост этого вида наблюдается в теплоумеренной зоне и переходной к холодноумеренной. Холодостоек, выносит сильные морозы (-25 °С), устойчив к грибковым заболеваниям. В Японии имеется около 100 сортов, среди которых самые крупноплодные каштаны в мире (масса одного плода около 80 г, диаметр 6 см), широко экспортируемые в США.
Китайский каштан мягчайший (С. mollissima) - дерево средней величины (высотой до 20 м), с белоопушенными снизу крупнозубчатыми листьями, шелковистоопушенной плюской и крупными орехами, диаметром 2 - 3 см. Распространен от Восточных Гималаев до Корейского полуострова и острова Тайвань. Китайцы длительное время культивируют этот вид, выведен ряд ценных крупноплодных сортов с высокими вкусовыми качествами. Во многих китайских городах, окруженных каштановыми насаждениями, продается большое количество плодов этого каштана, играющих важную роль в питании населения. Каштан мягчайший широко культивируется также в Корее, на Филиппинах, в Индии, Индокитае, Европе (Франция) и Северной Америке.
Кроме этого вида, в Китае встречаются еще 2 вида каштана: каштан Сегю (С. seguinii) - низкий кустарник (или небольшое дерево), распространенный в восточных и центральных районах на высоте 1000 - 1600 м над уровнем моря, и единственный представитель особой секции гипокастанон (Hypocastanon) - каштан Генри (С. henryi) - дерево высотой около 25 м, весьма обычное в горных широколиственных лесах центральных и западных провинций. У этого каштана мелкие, но съедобные плоды и очень хорошая древесина.
На юге США растет несколько видов кустарниковых каштанов, принадлежащих к особой секции баланокастанон (Balanocastanon) с одно-цветковыми женскими дихазиями. Плюска этих видов раскрывается 2 створками и содержит только 1 орех. От видов рода кастанопсис они отличаются только симметрией плюски и летне-зеленостью.
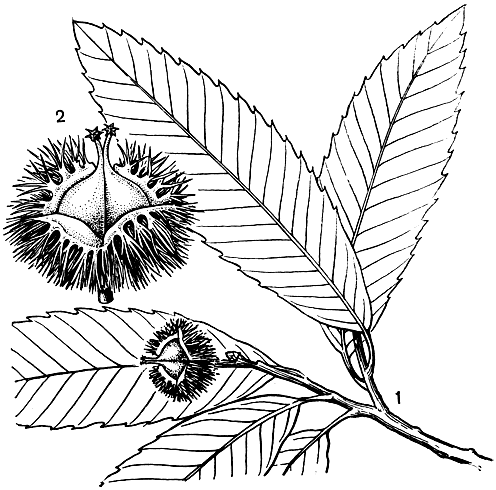
Рис. 158. Каштан зубчатый (Castanea dentata): 1 - побег с плодами; 2 - плюска с орехами
Каштан карликовый (С. pumila) - кустарник высотой 1 - 2 м, в благоприятных условиях достигающий размеров маленького дерева с беловато-войлочными снизу, грубозубчатыми, продолговато-эллиптическими листьями. Плоды этого вида размером с лесной орех имеют приятный вкус. Растет на востоке США от Пенсильвании до Флориды. Холодостойкий вид, устойчив к грибковым заболеваниям. Древесина крупных экземпляров прочная, тяжелая, не гниющая в контакте с водой, высоко ценится в США.
Каштан олъхолистный (С. alnifolia) - один из самых мелких представителей семейства, как правило, кустарник высотой до 30 см и даже в благоприятных условиях не превышает 1 м. Распространяется с помощью подземных столонов. Встречается на Атлантическом побережье США (Луизиана, Миссисипи, Алабама, Флорида, Джорджия, Южная Каролина) на песчаных холмах.
Точно так же как виды каштана - производные типовой секции рода кастанопсис, виды рода литокарпус (Lithocarpus, рис. 159) произошли непосредственно от его секции псевдо-пазания. Вечнозеленые деревья средних размеров, немногие из которых достигают значительной высоты (до 45 м), реже - кустарники высотой 1 - 3 м, обычно с глубоко идущим и разветвляющимся стержневым корнем. Около 250 видов литокарпусов распространены в тропических, субтропических, реже в умеренных широтах Гималаев, Восточной Азии и горных поясах Юго-Восточной Азии - от уровня моря до высоты 3000 м (от Непала до Корейского полуострова, Японии, острова Тайвань, Индокитая, Филиппин и Новой Гвинеи; один вид литокарпус густоцветковый - L. densiflora - на западе США). Многие виды являются важными компонентами горных дождевых дубово-лавровых лесов.
Листья литокарпусов толстые, кожистые,. цельнокрайние или мелкозубчатые, средних размеров, но иногда очень большие (у некоторых видов до 35 - 45 см), всегда с развитыми черешками, у крупнолистных форм иногда очень толстыми (до 8 мм). Цветки развиваются на жестких вертикальных или косо вверх торчащих специализированных побегах, выходящих из пазух верхних листьев вегетативных побегов. Имеются 4 типа соцветий - мужские (в нижней части побега), женские, обоеполые (с женскими цветками у основания, мужскими - у верхушки) и смешанные (мужские цветки располагаются поочередно на оси); у большинства видов наблюдается сочетание обоеполых (верхних) и мужских (нижних) соцветий. Мужские цветки одиночные или чаще собраны в 3 - 24-цветковые дихазиальные группы, с 6 - 12 тычинками на удлиненных нитях, пыльники прикреплены к нитям спинками, с рудиментарным гинецеем (иногда даже с 3 функционирующими столбиками). Женские дихазии одно-цветковые, являются продуктом редукции многоцветковых дихазиев кастанопсиса, но, в свою очередь, у многих видов они собраны в дихазиальные группы высшего порядка (по 2 - 24) и имеют соответствующие прицветники. Женские цветки похожи на таковые кастанопсиса - с 3-членным 2-круговым околоцветником, доли которого, в отличие от околоцветника кастанопсиса, не мягкие и мясистые, а кожистые, с 10 - 12 стаминодиями, 3 - 6-гнездной завязью ж 3 - 6 цилиндрическими столбиками, рыльца терминальные, точковидные. У некоторых малезийских видов стаминодии хорошо развиты.
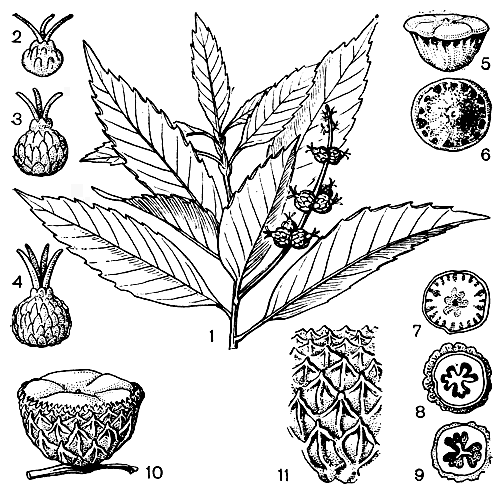
Рис. 159. Литокарпус полушаровидный (Lithocarpus hemisphaericus): 1 - побег с женским соцветием; 2 - 4 - женские цветки; 5 - 6 - желудь; 7 - 9 - поперечные разрезы желудя; 10 - желудь с плюской; 11 - чешуи плюски
Каждый женский цветок в конце цветения и завязь окружены собственной плюской, характерной особенностью которой, в отличие от плюски кастанопсиса, является отсутствие шовных зон - зона роста плюски у литокарпусов, как и у дубов, непрерывна по окружности. Иногда в дихазиальных группах развивается только 1 цветок, а остальные остаются недоразвитыми либо происходит частичное или полное срастание плюсок. В отличие от плюски кастанопсиса плюска литокарпуса никогда не бывает покрыта иглами, ветвистыми шипами или бугорками - на ней развиваются, как и на плюске дубов, чешуйки, свободные либо сросшиеся в кольца или спирали. Глубокая, закрывающая большую часть плода плюска, часто значительной толщины, с развитыми жесткими чешуйками выполняет защитную функцию, предохраняя от повреждений несозревшие желуди, а также функцию распространения плодов, цепляющихся своими часто оттопыренными чешуйками за шерсть животных. Плоды - желуди.
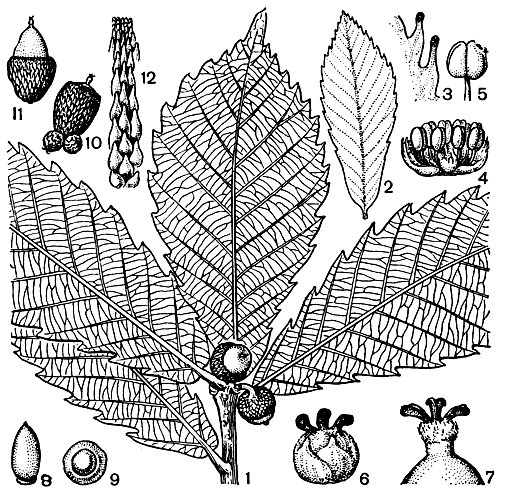
Рис. 160. Дуб Гриффита (Quercus griffithii): 1 - побег с плодами; 2 - молодой лист; 3 - край листа; 4 - мужской цветок; 5 - пыльник; 6 - женский цветок; 7 - верхушка завязи со столбиком; 8 - 9 - желуди; 10 - 11 - желуди с плюской; 12 - чешуи плюски
Цветение литокарпуса обычно происходит весной. В тропиках Малезии сезон цветения наступает непосредственно за самым сухим месяцем года: на Молуккских островах - в марте - апреле, на острове Ява - в июле - августе. Опыление у всех видов, как это видно по форме мелкого терминального точковидного рыльца, производится главным образом насекомыми - мелкими мухами, пчелами и жесткокрылыми, поедающими пыльцу. Созревание плодов у тропических видов происходит через 6 месяцев после цветения. Цветение и плодоношение у многих видов происходит ежегодно, у некоторых - весьма нерегулярно. Прорастание желудей подземное, и в благоприятных условиях температуры и влажности происходит в течение 1 - 2 недель после их опадания.
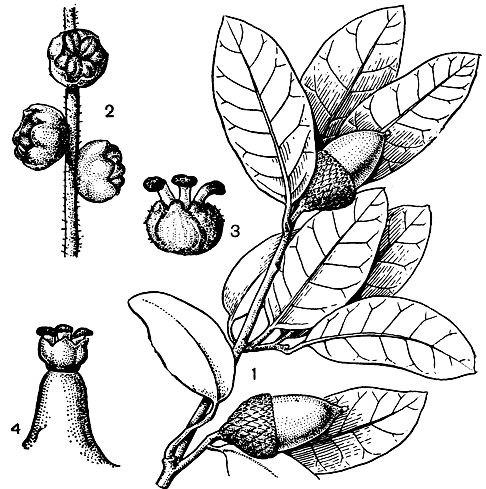
Рис. 161. Дуб вирджинский (Quercus virginiana): 1 - побег с желудями; 2 - мужские цветки; 3 - женский цветок; 4 - верхушка завязи с периантоподиумом
Большинство видов литокарпуса имеют крупные, тяжелые плоды, которые не могут разноситься ветром; в их распространении важную роль играют белки, для которых они являются существенным видом корма. В некоторых благоприятных для произрастания этих деревьев районах тропической Азии, где нет белок (например, в горных лесах острова Суматра выше 1500 м), литокарпус не встречается. Некоторое значение в распространении плодов играют птицы, переносящие их иногда на значительное расстояние, а также реки - слегка подсохшие плоды какое-то время не тонут в воде.
Древесину литокарпуса используют для строительства домов, изготовления мебели, повозок, деталей рисовых мельниц, для всякого рода столярных работ, для железнодорожных шпал. Некоторые виды дают очень прочную, твердую древесину, но в условиях тропиков она легко повреждается термитами, древесина лишь немногих видов устойчива к этим насекомым. Древесина единственного калифорнийского вида - литокарпуса густоцветкового, хотя и твердая, но ломкая и используется главным образом на топливо. Кора многих видов содержит много танинов. Танины имеются в значительных количествах и в плюсках. Желуди богаты крахмалом и растительными маслами, и после высушивания, когда они в значительной степени теряют вяжущий привкус, их использует в пищу местное население (в Японии, Китае, Индокитае и Малезии).
Род дуб (Quercus, рис. 160, 161, 162, табл. 41) с эволюционной точки зрения является анемофильным производным рода литокарпус и очень близок к нему; оба рода долгое время даже систематиками не различались, как таковые. Различие между этими родами заключается в разной форме рылец.
Дубы (вероятно, около 450 видов) являются важнейшими лесообразующими древесными породами умеренных широт и горных поясов северного полушария, а также компонентами тропического и субтропического (дождевого и сухого) леса Юго-Восточной Азии (карта 15). Наиболее богаты видами дубов Северная и Центральная Америка, где они распространены от юго-востока Канады до Колумбийских Анд; Европа, Северная Африка и Юго-Западная Азия бедны видами этого рода; Сибирь и Центральная Азия вообще не имеют аборигенных видов, и только Восточная и Юго-Восточная Азия представляют сравнимый по видовому богатству с американским центр разнообразия рода. Северная граница ареала дуба в Евразии находится на широте около 63° в Скандинавии, южная уходит, как и в Южной Америке, на несколько градусов за экватор (острова Суматра, Ява).
Большинство дубов - высокие деревья (25 - 30 м), на равнинах обычно выше, чем в горах; отдельные экземпляры достигают громадной высоты (до 55 м) и имеют в возрасте 700 - 900 лет толстые (диаметром до нескольких метров) стволы. Немногие виды являются низкими кустарниками и даже кустарничками, покрывающими почву ковром высотой около 0,5 м и в самых благоприятных условиях редко достигающими в высоту 2 - 3 м (дуб кустарниковый - Q. fruticosa - в Испании). Многие виды, обычно являющиеся большими деревьями, в высокогорьях, в сухих местах или при частых рубках и травле скотом не вырастают выше кустарника.
Корневая система дубов обычно мощная, стержневого типа (часто до 5 м в глубину), однако при наличии подстилающего известнякового горизонта или других плотных пород, на подзолистых или переувлажненных почвах становится поверхностной. Некоторые виды дубов тропического дождевого леса у основания ствола развивают мощные досковидные корни.
Особенности коры (точнее, перидермы) часто являются характерными видовыми признаками: у одних видов она гладкая или долгое время остается таковой и несет лишь чечевички, у большинства видов покрыта трещинами различной толщины или разбита на прямоугольные пластинки. Например, у дуба обыкновенного, или черешчатого (Q. robur), кора с хорошо выраженными продольными глубокими извилистыми трещинами и короткими поперечными щелями, углубляющимися с возрастом; у дуба скального (Q. petraea) она разделена на отдельные пластинки. У дуба пробкового (Q. suber), в меньшей степени у дуба изменчивого (Q. variabilis) сильно развит пробковый слой коры; у американского дуба крупноплодного (Q. macrocarpa) пробковый слой образует характерные "крылья" на ветвях.
Ветвление у дубов, особенно хорошо видное в зимний период, кажется очень резким и угловатым; это происходит от частой смены направления роста побегов, обусловленной большим светолюбием дубов и стремлением дерева развивать листья только на освещенной части крон. У дубов на концах побегов развиваются настоящие верхушечные почки (чего нет, например, у кастанопсисов); возможный при этом моноподиальный рост побега практически осуществляется очень редко в связи с частым повреждением верхушечных почек и "перевершиниванием" побега за счет развития боковых почек. При наличии нескольких периодов нарастания побега в один сезон такая смена может происходить несколько раз в течение года. У большинства видов почки пятигранные, с коричневыми чешуйками, расположенными в 5 рядов. У основания почек часто развиты 2 узких прилистника.
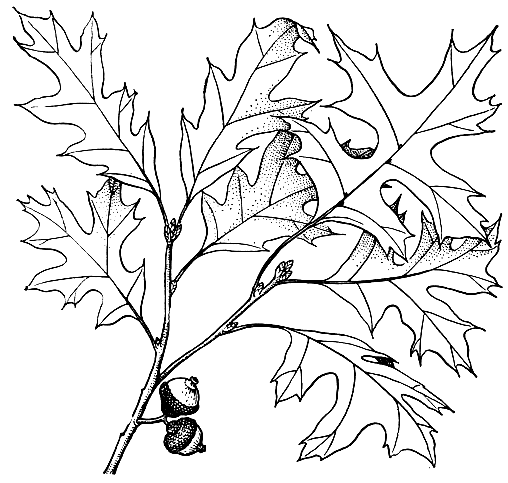
Рис. 162. Побег с желудями дуба болотного (Quercus palustris)
Листья у дубов в среднем длиной 7 - 15 см (у вечнозеленых видов они обычно меньше). В редких случаях, как правило у листопадных видов, листья достигают гигантских размеров - в длину до 40 см и в ширину до 30 см (дуб зубчатый - Q. dentata, дуб крупнолистный - Q. macrophylla), у ряда вечнозеленых видов листья чрезвычайно мелкие. Черешки обычно короткие, у некоторых видов почти полностью редуцированы, у весьма немногих видов достигают в длину 7 см. В роде существует несколько морфологических типов листьев: 1) тип лавра - вечнозеленые, цельнокрайние (или только у верхушки мелкозубчатые), с равноотстоящими параллельными жилками (почти весь подрод циклобаланопсис - Cyclobalanopsis и ряд видов подрода эритробаланус - Erythrobalanus); 2) тип падуба - вечнозеленые, жесткие, мелкие, с немногочисленными жилками, иногда раздваивающимися на конце, и колючими зубцами (подрод гетеробаланус - Heterobalanus); 3) тип каштана - летнезеленые листья с многочисленными параллельными равноотстоящими друг от друга вторичными жилками, идущими обычно в заостренные (иногда с длинными остроконечиями) зубцы (подрод Quercus); 4) тип лопастных или лопастно-зубчатых листьев с равномерно или неравномерно расположенными боковыми жилками (подрод Quercus); 5) тип лопастно-зубчатых листьев, лопасти которых длинно заострены на концах, с неравноотстоящими друг от друга боковыми жилками и многочисленными интеркалярными (подрод эритробаланус). Форма листьев может быть от почти линейной до округлой, основание - от выемчатого до узкоклиновидного, верхушка - от длинновытянуто-заостренной до округлой. Часто у одного и того же экземпляра нижние листья побега сильно отличаются по форме и размерам от верхних (гетерофиллия); это почти всегда имеет место у видов, подверженных перерыву вегетации не только зимой, но и летом вследствие высоких температур и недостатка влаги.
Плюска у дубов развивается в конце цветения и представляет собой полный гомолог подобной структуры литокарпуса, от которой она не отличается ни внешним видом, ни анатомическим строением. Как у литокарпуса и некоторых кастанопсисов, плюска дубов окружает редуцированный (1-цветковый) дихазий.
В очень редких случаях у некоторых видов подрода циклобаланопсис наблюдается появление в плюске не одного, а нескольких (обычно двух) желудей.
Форма плюски у дубов очень разнообразна. Она покрывает иногда до 2/3 желудя, но, в отличие от плюски литокарпуса, никогда не покрывает плод целиком и никогда полностью не срастается с ним, как это происходит у многих видов последнего рода. Диаметр плюски варьирует от 5 - 6 мм до 5 - 6 см. На наружной поверхности она покрыта чешуйками - прижатыми или оттопыренными, разнообразной величины и формы, свободными или (подрод циклобаланопсис) полностью сросшимися в кольца (но не в спирали, как иногда бывает у литокарпуса).
Цветение дуба обыкновенного происходит в средней полосе в конце апреля - в мае, одновременно с распусканием листьев. После раскрытия мужских цветков пыльца остается жизнеспособной около 5 суток; пылению благоприятствуют повышенная температура и ветер; пыление уменьшается или вообще прекращается при моросящем дожде (пыльники часто не раскрываются) и во время заморозков может начаться массовое опадение соцветий. Опыление у крупных деревьев обычно продолжается несколько дней, но у некоторых американских кустарниковых дубов в благоприятных погодных условиях заканчивается за несколько часов. У дубов часто наблюдается протандрия - обычно пыльники уже сбросили пыльцу, когда рыльца женских цветков того же побега еще совсем слабо развиты.
В момент опыления завязь еще не сформирована и ее полость представляет собой трехлучевую щель. Семязачатки у большинства видов с одногодичным созреванием желудей заканчивают развитие только через 1 - 2 месяца после опыления, когда желудь станет величиной с горошину. У видов с двухгодичным созреванием плодов оплодотворение происходит только через год после опыления.
Рост и формирование желудей у дуба обыкновенного происходит с начала августа по сентябрь. До середины августа мясистые семядоли заполнены главным образом сахаром, а впоследствии появляются крахмальные зерна и капли жира. Со второй половины сентября по ноябрь происходит созревание и опадение желудей.
Плюска дубов, как и литокарпусов, имеет вполне определенную функцию - защиту основания растущего желудя, где перикарпий еще очень мягкий. Она развивается быстрее желудя и вначале полностью окружает плод7! так что только околоцветник и столбики торчат на его верхушке.
Распространение желудей производится ветром (если он достаточно силен), но главным образом животными, а также водой. Птицы (сойки, голуби, фазаны) и грызуны иногда довольно далеко разносят желуди, используя их в качестве пищи и делая запасы. Белки и полевые мыши очень любят желуди и поедают их в больших количествах, а также разносят их по своим норам - часть запасов остается неиспользованной и может прорасти. В горных районах желуди переносятся также реками и ручьями. Желуди дубов подрода дуб не требуют периода покоя и прорастают осенью, зимуя под снегом в проростках, иногда с большими потерями в суровые зимы; виды подрода эритробаланус требуют для прорастания периода покоя и прорастают только весной. Из-за обилия питательных резервов в семядолях прорастание происходит быстро. Околоплодник раскалывается на верхушке, откуда сначала появляется корешок, направляющийся вертикально вниз. Стебель, развивающийся вслед за корешком, несет у основания несколько очень мелких чешуйчатых листьев; первые настоящие зеленые листья обычно очень короткие и отличаются от взрослых листьев супротивным расположением, меньшей рассеченностью и меньшим числом боковых жилок. Только у подрода эритробаланус первые листья не обнаруживают резких отличий от взрослых листьев.
В первые годы жизни молодое растение требует для нормального развития некоторого затенения, что обычно имеет место при естественном возобновлении в лесу. Это всегда учитывают при лесопосадках: желуди сажают в заранее подготовленные полосы из кустарников (лещина, желтая акация, вяз и др.) или даже между рядами кукурузы (растения притеняют молодые дубки и одновременно служат им подгоном) либо применяют метод густых культур, где происходит самозатенение. На открытых местах молодые растения не выдерживают конкуренции со злаками; последние могут погубить даже 5-метровые деревья при посадках в степных районах.
Кроме семенного размножения, дубы обладают способностью давать обильную прикорневую поросль и прикорневые отводки; очень редко, только при повреждении поверхностных корней, появляются корневые отпрыски. Большинство видов дубов легко скрещивается между собой и дает плодовитые гибриды.
Подрод циклобаланопсис является одним из; самых древних в роде; его виды произошли, по-видимому, от секции циклобаланус рода литокарпус и характеризуются кольчатыми плюсками. Виды этого подрода растут исключительно в Восточной и Юго-Восточной Азии (от Западных Гималаев до Японии, от Центрального Китая до островов Суматра и Ява); около 10 видов растут уже в южном полушарии. Виды, растущие в тропиках и являющиеся компонентами тропического и субтропического горного дождевого леса, обычно имеют тонкие, мягкие листья; виды муссонного климата, способные переносить длительную зимнюю засуху, имеют толстые, жесткие листья, иногда с трабекуло видно выступающими снизу жилками, обычно густо опушенные, некоторые индокитайские виды даже сбрасывают в сухой период листья. Большинство видов не поднимается в горы выше 2000 м и лишь немногие виды в Гималаях, в Китае и во Вьетнаме растут в зоне умеренного климата выше 2000 м над уровнем моря.
Виды подрода гетеробаланус имеют обычно более или менее мелкие, жесткие, колючезубчатые или цельнокрайние листья, боковые жилки которых часто вильчато ветвятся на концах. Это жестколистные вечнозеленые деревья и кустарники, распространенные в умеренных (редко в субтропических) поясах гор Средиземноморья, Юго-Западной, Восточной и Юго-Восточной Азии, Северной и Центральной Америки. В Гималаях некоторые виды этого подрода являются огромными деревьями и образуют чистые или смешанные леса, другие, особенно в горных районах Китая и Индокитая, - небольшими деревьями или кустарниками, формирующими редколесья на верхней границе распространения древесной растительности в горах или маквисоподобные формации, характерные для гор Центрального и Западного Китая. В Средиземноморье некоторые виды этого подрода, в том числе дуб пробковый, иногда образуют чистые или смешанные леса, по генетическому составу входящих в них видов дубов сходные с таковыми Восточной Азии, но чаще являются важными компонентами маквиса вместе с земляничным деревом, вереском, можжевельником, фисташкой, маслиной, филлиреей и другими растениями. Аналогичная формация (чапараль) встречается в юго-западных районах Северной Америки (в сухих районах от Орегона до Нью-Мексики). Множество видов этой группы обитает в сухих районах Скалистых гор, южных атлантических штатах США ("живые дубы"), на островах Карибского моря и в горах Мексики.
Подрод эритробаланус является одним из наиболее древних в роде. Многочисленные виды (около 200) этого рода имеют летнезеленые цельнокрайние или зубчатые листья с длинными остроконечиями на зубцах, осенью приобретающие красивую красную окраску (откуда название - красные дубы). Плюска красных дубов имеет широкие плоские чешуйки, желуди созревают на второй год после опыления и на верхушке имеют столбчатый вырост. Это исключительно американские виды, распространенные в атлантических и западных штатах США, а также на равнинах и в горах Центральной Америки, где они особенно многочисленны. Многие виды этой группы являются мезофитами и представляют собой важнейшие в хозяйственном отношении лесообразующие породы, растущие в самых разнообразных условиях: в поймах и долинах рек, в болотах {дуб болотный - Q. palustris, рис. 162), в предгорьях и горах востока США от северных штатов до Техаса и долины реки Миссисипи, а также в умеренных поясах гор Центральной Америки (Гватемала, Гондурас, Коста-Рика). Более ксерофильные, обычно цельнолистные виды растут в сухих районах юга и запада США и Центральной Америки, обычно в виде кустарников, иногда образующих непроходимые заросли. Красные дубы менее требовательны к почве, чем виды других групп, хотя в общем являются кальцефобами, и представляют, несмотря на свою древность, весьма агрессивную группу, вполне конкурентоспособную не только с другими дубами, но и с хвойными, которых во многих случаях успешно вытесняют. Они растут и возобновляются быстрее "белых" дубов (подрод Qnercus), устойчивы к низовым пожарам; их горькие желуди почти не поедаются животными, и они меньше страдают от широко распространенных среди "белых" дубов грибковых заболеваний.
Виды подрода Quercus являются наиболее эволюционно продвинутыми в роде. Это листопадные зубчато - или лопастнолистные виды (иногда вторично цельнокрайние), с одно-, редко двухгодичным созреванием желудей, распространенные на трех континентах - в Восточной и муссонных областях Юго-Восточной Азии, Европе и Северной Африке, Северной и Центральной Америке, где в большинстве случаев являются важными лесообразующими породами или содомин антами в смешанных широколиственных лесах. В Северной Америке на равнинах и в горах восточных штатов и в Северной Мексике они вместе с красными дубами образуют дубово-кариевые, дубово-лириоденд-роновые и смешанные с хвойными (часто дубово-сосновые) леса. Несколько видов секции макробаланус (Macrobalanus) живет в Мексике в горах Сьерра-Мадре, в горах Гватемалы, Гондураса, Панамы и Коста-Рики.
В Европе дубы растут повсюду - от побережья Средиземного моря до 63° северной широты в Скандинавии, но в настоящее время ввиду уничтожения их на равнинах преобладают в горах (главным образом до высоты 1500 - 2000 м), где образуют чистые или дубово-грабовые, дубово-каштановые, дубово-буковые леса, а также леса с примесью липы, клена, ильма, ясеня и других пород; на севере растут в зоне хвойных лесов. Здесь 4 важнейших вида - дуб обыкновенный, или черешчатый, дуб скальный (Q. petraea), дуб пушистый (Q. pubescens) и дуб австрийский (Q. cerris) - дают основную массу поделочной и строевой древесины. В Восточной и Юго-Восточной Азии дубы этого подрода занимают умеренные пояса гор, выходя на севере на равнины (дуб монгольский - Q. mongolica). В редких случаях дубы этого подрода спускаются до субтропического и даже тропического пояса (обычно на вторичных местообитаниях только в районах с сильной зимней засухой), оставаясь при этом типично листопадными деревьями.
Наибольшее практическое использование находит древесина дуба, которая является важнейшим лесопильным материалом, применяемым для наиболее ответственных работ, где нужны прочность, твердость, упругость, стойкость к факторам внешней среды, - в судостроении, авиационной промышленности, транспорте, строительстве жилищ, разного рода столярных работах и т. д. Кроме того, древесина почти всех видов дубов дает прекрасное топливо, калорийность которого растет вместе с плотностью; дубовый древесный уголь отличается хорошим качеством. Наиболее твердую древесину дают виды подрода гетеробаланус, очень высокими эксплуатационными показателями характеризуется и древесина видов подрода Quercus. Древесина красных американских дубов отличается от прочих розоватым или красноватым оттенком, она более мягкая. Однако в этом подроде также имеются виды с весьма ценной древесиной. Древесина видов подрода циклобаланопсис в среднем по эксплуатационным показателям гораздо хуже таковой видов умеренных областей - не очень твердая, колкая, часто коробится и повреждается насекомыми. Наилучшую древесину из циклобаланопсисов дают некоторые гималайские и один индокитайский (Q. helferiana) виды.
Кора, древесина, листья и плюски, а также галлы, образующиеся на листьях, очень богаты танинами, используемыми для выделки кож. Особенно часто применяют для этой цели кору и плюски дубов. Ценность этих видов сырья повышается в южных районах; богатство танинами уменьшается с возрастом дерева - наилучший выход танинов получают от деревьев в возрасте 15 - 35 лет. Дуб пробковый и дуб изменчивый дают пробку промышленного значения, используемую в холодильной промышленности, моторостроении, судостроении, медицине и в быту. Наилучшую пробку получают из дуба пробкового, дикорастущего и широко культивируемого для этой цели в странах Западного Средиземноморья. Наибольший выход пробки получают от марокканских деревьев, которые часто вдвое выше европейских и, кроме того, дают съедобные, похожие на каштаны желуди. Плантации пробкового дуба на Черноморском побережье Кавказа занимают около 200 га и далеко не удовлетворяют потребности страны в этом виде сырья. Другой пробконос - дуб изменчивый - имеет промышленное значение в Китае, где растет в большинстве провинций. В настоящее время этот более холодостойкий вид интродуцирован на Кавказ (Майкоп, Красная Поляна, Геленджик, Анапа) и является перспективным для разведения в среднегорной зоне.
Желуди дубов играли важную роль в питании людей в доисторические эпохи, и даже в более поздние времена горькие желуди ряда видов использовались для выпечки хлеба, особенно в неурожайные годы. У некоторых средиземноморских дубов желуди не только вполне съедобны, но и приятны на вкус; в ряде стран (Португалия, Испания, Италия, Греция, Турция, Алжир) они широко используются в пищу местным населением. Из американских дубов только белые (секция принус - Prinus) имеют съедобные и приятные на вкус желуди; в настоящее время некоторые индейские племена иногда используют в пищу плоды некоторых видов этой секции. Некоторые виды подрода циклобаланопсис также дают съедобные желуди: в Японии местное население собирает плоды дуба сизого (Q. glauca). Из желудей своего природного пробконоса дуба изменчивого китайцы приготовляют национальное блюдо "дофун". Желуди часто служат в качестве суррогата кофе, или ими фальсифицируют натуральный; сладкие желуди средиземноморских видов, растертые в виде пудры, добавляют в шоколад или в муку. Муку сладких желудей, смешанную с какао, используют в Испании против энтерита.
Для многих зверей (кабанов, оленей, медведей) и птиц (соек, голубей, фазанов) желуди являются прекрасным кормом; в урожайные годы их вместо кукурузы добавляют в корм домашним животным и птицам. Листья дуба иногда применяют в качестве корма для скота, однако молодые побеги весьма токсичны для животных и поедание их грозит смертью. В Китае листья многих видов используют для выкармливания личинок шелковичного червя и как удобрение (в смеси с глиной).
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'