Строение семязачатков
Основные черты строения семязачатков мы уже рассматривали в предыдущем томе "Жизни растений". Но напомним еще раз, что семязачаток состоит из двух основных частей - мегаспорангия (нуцеллуса) и окружающего его покрова, называемого интегументом. Интегумент не полностью закрывает мегаспорангий, на верхушке семязачатка его края не срастаются полностью и оставляют отверстие в виде канала для внедрения пыльцевых трубок. Это микропиле, или, иначе, семявход. Нижняя часть семязачатка, т. е. место ее перехода в семяножку (фуникулус), называется халазой. Некоторые авторы, в том числе даже такой известный эмбриолог, как индийский ботаник П. Махешвари (1950), весь семязачаток приравнивают к мегаспорангию. Это большая ошибка. Семязачаток - это не только мегаспорангий, это мегаспорангий плюс интегумент.
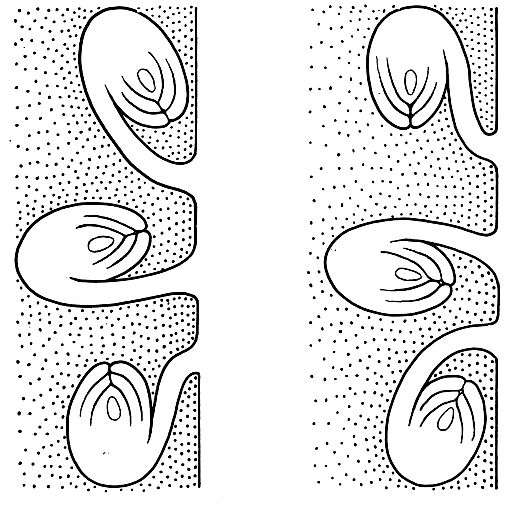
Рис. 27. Типы анатропного семязачатка в зависимости от его поворота в гнезде и от направления фуникулуса: в левом ряду - эпитропные, в правом - апотропные
Читатель может спросить: почему мегаспорангий семязачатка называют нуцеллусом, если известно, что это мегаспорангий? Дело в том, что долгое время это не было известно, пока в 1851 г. великий немецкий ботаник В. Гофмейстер не установил, что образование, давно известное под названием нуцеллуса, представляет собой не что иное, как мегаспорангий. Но, как часто бывает в науке, даже после замечательных исследований Гофмейстера мегаспорангий семенных растений продолжали называть нуцеллусом, а некоторые ботаники, как, например, американский морфолог А. Имс (1961), подвергали сомнению даже само наличие мегаспорангия у цветковых растений.
Однако большинство ботаников справедливо считают, что наличие у цветковых мегаспорангия, правда очень упрощенного и редуцированного, не вызывает сомнений. Ведь в нуцеллусе имеются два необходимых элемента каждого спорангия - спорогенная ткань (обычно крайне редуцированная) и стенка, которая, однако, гистологически сильно упрощена. Совершенно очевидно, что нуцеллус цветковых морфологически вполне соответствует нуцеллусу голосеменных, т. е. они представляют собой гомологичные образования и являются мегаспорангиями.
Как и у голосеменных, мегаспорангий цветковых окружен интегументом. Однако, в то время как у голосеменных имеется лишь один интегумент, который, согласно гипотезе Маргариты Бенсон (1904), произошел из обеспложенного периферического слоя спорангиев, окружавших центральный фертильный спорангий, у подавляющего большинства цветковых имеются два интегумента. В связи с этим различают битегмальные (от лат. bis - дважды и tegmen - покров) семязачатки, т. е. семязачатки с двумя интегументами, и унитегмальные ( от лат. unus - один), т. е. семязачатки с одним интегументом.
В разных линиях эволюции цветковых растений из битегмальных семязачатков произошли унитегмальные. Унитегмальные семязачатки характерны для березовых, мириковых, ореховых, вересковых и близких им семейств, кизиловых, аралиевых, зонтичных, для различных семейств двудольных, характеризующихся спайнолепестным венчиком, и для некоторых однодольных. По-видимому, в большинстве случаев унитегмальные семязачатки произошли из битегмальных в результате слияния обоих покровов, но в ряде случаев они возникли вследствие недоразвития внешнего или чаще внутреннего покрова. Так, в семействе раффлезиевых наблюдается редукция внешнего интегумента, а у рода митрастемон (Mitrastemon) он совершенно подавлен и семязачатки унитегмальные. У родственного семейства гидноровых семязачаток постоянно унитегмальный. В то же время у некоторых представителей семейства розовых, таких, как гравилат, земляника, лапчатка и манжетка, унитегмальный семязачаток возник в результате недоразвития внутреннего покрова. В некоторых семействах, таких, как лютиковые, розовые, бобовые, ивовые, пальмы, орхидные и другие, даже близкие роды отличаются числом интегументов. Это показывает, что унитегмальные семязачатки возникли из битегмальных независимо в самых разных ветвях развития цветковых растений.
У некоторых паразитных двудольных покров семязачатка исчезает полностью и мегаспорангий оказывается, таким образом, голым. Такие атегмальные (от греч. а - частица отрицания) семязачатки характерны для многих представителей порядка санталовых. У некоторых баланофоровых утрачен не только покров семязачатка, но даже стенка мегаспорангия.
Но откуда появился второй интегумент у битегмальных семязачатков? Этот вопрос оказался одним из наиболее трудных в морфологии растений, и его еще нельзя считать вполне решенным. По всей вероятности, из двух покровов лишь внутренний является настоящим интегументом, соответствующим единственному интегументу голосеменных. Что же касается внешнего интегумента, то скорее всего он имеет другое происхождение. Ряд авторов высказал предположение, что внешний интегумент цветковых растений имеет не спорангиальное, а листовое происхождение. По мнению этих авторов, внешний интегумент произошел из купулы ("плюски") семенных папоротников - этих вероятных предков цветковых растений. В видоизмененной форме купула сохранилась и у некоторых примитивных голосеменных. Так, антлийский палеоботаник Мэри Стоупс (1905) рассматривала наружный мясистый слой семени ныне живущих саговниковых, или саркотесту, как образование, гомологичное купуле типа лагеностомы (Lagenostoma). Но из купулы произошел не только внешний слой покрова семязачатка ряда голосеменных, но, по-видимому, и внешний интегумент цветковых растений. Идея купулярной природы внешнего интегумента цветковых растений была выдвинута независимо французским ботаником А. Госсеном (1946) и английским ботаником Дж. Уолтоном (1953), а сравнительно недавно - американским ботаником Дж. Л. Стеббинсом (1974). Она представляется вполне убедительной.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'