Порядок Селагинелловые - Selaginellales
Познакомимся с большим родом Селагинелла (Selaginelia), который охватывает около 700 видов, распространенных преимущественно в тропических странах. В нашей флоре известно несколько видов селагинеллы, имеющих сравнительно небольшой ареал. В оранжереях для создания газонов культивируют ряд тропических видов, некоторые из них можно выращивать в комнатных тепличках, что дает возможность иметь на учебных занятиях живой материал.
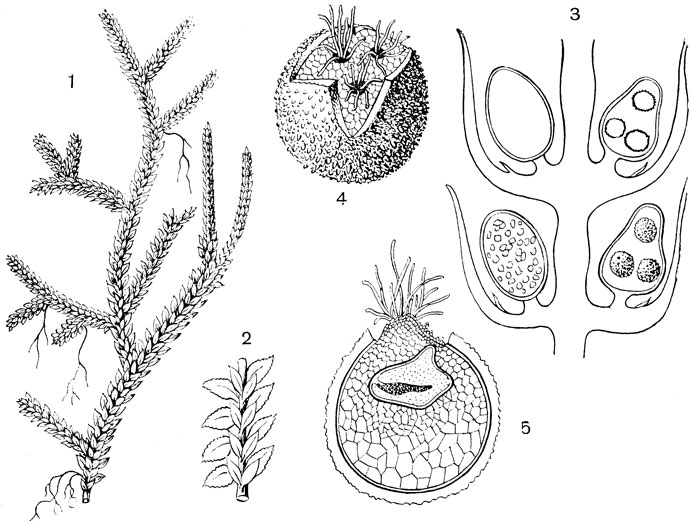
Рис. 60. Селагинелла (Selaginella): 1 - общий вид растения с колосками; 2 - формы листьев и расположение их; 3 - продольный разрез колоска с мегаспорангиями и микроспорангиями; 4 - женский заросток, окруженный оболочкой мегаспоры; 5 - продольный разрез заростка с зародышем спорофита
Внешнее строение. Виды селагинеллы большей частью нежные травянистые растения со стелющимися или слабо восходящими побегами, покрытыми мелкими зелеными листочками. У нашего северного вида селагинеллы плауновидной (Selaginella selaginoides) все листья одинаковые, спирально расположенные; у большинства же оранжерейных видов листья разные, расположенные в четыре ряда: два ряда сравнительно крупных боковых листьев и два ряда чередующихся, более мелких листьев на верхней (спинной) стороне побега (рис. 60, 1, 2). Ветвление более или менее явственно дихотомическое, причем ветви нередко располагаются в одной плоскости. У основания листовой пластинки всех видов селагинелл имеется крохотная пленчатая чешуйка, называемая язычком, функция которой не выяснена. У многих видов корни отходят не непосредственно от стебля, а от особых стеблевидных, но безлистных, свешивающихся вниз выростов, называемых корненосцами (ризофорами).
Размножение. Спороносные колоски селагинеллы сидят на концах вегетативных побегов без подставок. У Selaginella selaginoides колоски одиночные, овально-цилиндрические, со спирально расположенными спорофиллами; у большинства оранжерейных видов колоски многочисленные, призматические (четырехгранные), с накрест сидящими супротивными спорофиллами.
В колосках всех видов селагинеллы на одинаковых спорофиллах находятся разные спорангии: в верхней части колоска развиваются большей частью красноватые микроспорангии, в нижней асти - преимущественно бледно-желтые мегаспорангии, содер-ащие только четыре (иногда и меньше) мегаспоры, возникшие з одной материнской клетки (тетрада) (рис. 60, 3). Выделив из колоска микро- и мегаспорангии, вскроем их. Наблюдая микроспоры при большом увеличении, мы находим в них большое сходство со спорами равноспорового плауна (размер, форма, рисунок оболочки и пр.).
Затем рассмотрим в лупу мегаспорангий. Крупные выпуклости на его поверхности говорят о том, что споры в нем огромные, в чем мы убеждаемся, когда вскроем мегаспорангий и извлечем тетраду спор. В поле зрения микроскопа эти споры можно рассматривать лишь в отраженном свете. Сравнивая размеры микро- и мегаспор в одном поле зрения, найдем, что поперечник мегаспор в 15 - 20 раз больше поперечника микроспор.
Споры, выпавшие из спорангиев на влажную почву, прорастают; из микроспор формируются микроскопические мужские заростки, состоящие из примитивного антеридия и единственной вегетативной клетки, оболочка которой слегка выпячивается, подобно ризоиду. Развитие женского заростка протекает вначале внутри оболочки мегаспоры, которая позднее разрывается трехлучевой трещиной, при этом обнаруживается мелкоклеточная ткань женского заростка. На обнаженной части заростка появляются вскоре пучки ризоидов и несколько архегониев, погруженных в . ткань заростка (рис. 60, 4, 5). Иногда мегаспора начинает развиваться в женский заросток, находясь еще в мегаспорангии, и лишь позднее падает на почву. Оплодотворение возможно при наличии хотя бы капли воды, в которой одновременно окажутся и мужские и женские заростки.
Из зиготы развивается зародыш. Одна из крупных клеток у основания зародыша вытягивается в длину, продвигаясь в глубь заростка; эта клетка называется подвеском, черпающим питательные вещества, в начале развития зародыша, который, озеленяясь, переходит к самостоятельному питанию.
Закончив на этом изучение строения и циклов развития высших oспоровых растений, сделаем некоторые обобщения и выводы. Мы видели, что гаметофит всегда возникает из споры. Основная функция гаметофита - половое воспроизведение - осуществляется непременно в водной среде. Таков отпечаток, сохранившийся у гаметофита высших споровых от их далеких предков - водорослей - обитателей водной среды. Спорофит же всегда возникает заново только в результате полового процесса. Поэтому его способность изменять свою организацию соответственно условиям обитания значительно большая, чем у гаметофита, развивающегося из споры. Однако мохообразные и папоротникообразные не в одинаковой степени реализуют приспособительную способность своих спорофитов в воздушно-наземных условиях существования. Спорофит первых (спорогон), физиологически зависимый от гаметофита в течение всего периода своего существования, использует эту способность лишь в рамках присущей ему репродуктивной функции, в) органах образования и распространения спор. У спорофита же папоротникообразных, рано, на первых порах жизни, освобождающегося от такой зависимости и ведущего далее совершенно самостоятельный образ жизни, приспособительные способности выражаются значительно шире, в развитии не только репродуктивных, но и сложных вегетативных органов - корня, стебля и листа со свойственной им внутренней структурой. Благодаря этой организации папоротникообразные и сыграли видную роль в эволюции растительного мира. Тем не менее неразрывная генетическая взаимообусловленность спорофита, приспособленного к воздушно-наземным условиям, и гаметофита, тесно связанного с водной средой, привязывает папоротникообразные, подобно мохообразным, к избыточно влажным и тенистым местообитаниям. Экологическая обусловленность гаметофита препятствовала гигантским папоротникообразным завоевать сушу в период их расцвета на Земле. Только путем постепенного освобождения процесса полового воспроизведения от необходимости водной среды в момент оплодотворения высшие растения приобрели возможность расширить пределы распространения на более засушливые области Земли. Это достижение связано с появлением на Земле семенных растений, к изучению которых мы и переходим.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'