Порядок Аурикуляриевые (Auriculariales)
Характерный признак всех аурикуляриевых грибов - длинная цилиндрическая базидия с тремя поперечными перегородками. У некоторых представителей этой группы встречается хорошо развитый пробазидий, который после вырастания базидии сохраняется в виде гипобазидия. Различные представители этого порядка имеют совершенно отличные друг от друга плодовые тела: широко распростертые кортициевидные, сухие или студенистые либо студенистые с хорошо развитой уховидной шляпкой, мясистые, булавовидные, как у рогатиковых грибов, и, наконец, замкнутые, как у гастеромицетов. Все это свидетельствует о том, что в процессе приспособления к условиям среды, в результате конвергентной эволюции возникали в отдаленных друг от друга группах грибов очень схожие плодовые тела. Такие тенденции параллельной эволюции наблюдаются во всех трех порядках, которые раньше объединялись в искусственную группу гетеробазидиальных грибов.
Среди аурикуляриевых грибов встречаются как сапротрофы, так и паразиты на растениях, насекомых и других грибах.
Порядок аурикуляриевые делится на три семейства: септобазидиевые (Septobasidiaceae), аурикуляриевые (Auriculariaceae) и флеогеновые (Phleogenaceae), из которых самым древним и своеобразным можно считать септобазидиевые.
Семейство Септобазидиевые (Septobasidiaceae)
Характерный признак этого семейства - наличие толстостенных гипобазидий, которые, прежде чем дать начало четырехклеточным базидиям, проходят период покоя. Семейство охватывает около 200 видов, распределенных между двумя родами: септобазидиум (Septobasidium) и урединелла (Uredinella).
Род септобазидиум - обширная группа, в ней более 180 видов. Эти грибы растут на живых растениях, пораженных щитовкой. Их плодовые тела широкораспростертые, пленчатые, встречаются на ветвях и листьях растений, а под ними всегда можно найти щитовок, на которых гриб паразитирует, хотя некоторые насекомые остаются непораженными. Первые описания септобазидиума относятся к концу XVIII в. Но в то время эти своеобразные грибы были описаны как лишайники или как телефоровые грибы, и лишь в 1907 г. Генел и Литшауер открыли, что эти грибы всегда связаны со щитовками. Дальнейшие исследования показали, что гриб и насекомое связаны между собой сложными мутуалистическими отношениями. Большинство грибов этого рода растет в тропиках и субтропиках, в умеренном поясе встречаются немногие виды. Особенности биологии этого рода были подробно изучены американским микологом Коучем на септовазидиуме Бурта (Septobasidium burtii), который растет на разных видах дуба, а также на плодовых деревьях в южной части США и связан со щитовкой Aspidiotus osborni.
Плодовые тела септобазидиума Бурта резупинатные, округлые, диаметром до 15 см, многолетние, с хорошо заметными концентрическими зонами ежегодного роста, шириной 2-3 мм. Плодовое тело в разрезе толщиной до 2 см, состоит оно из трех слоев. Прямо на субстрате лежит тонкий субикулум, из которого вырастают радиальные, анастомозирующие стенки, образующие сложную систему туннелей и камер и поддерживающие верхний твердый бумаговидный слой - крышу. На самой поверхности плодового тела развивается гимений.
Под защищающей крышей гриба, в его сложном лабиринте, живут щитовки, некоторые из них остаются здоровыми на протяжении всей жизни, другие заражаются грибом. Здоровые и зараженные насекомые хорошо различаются: больных меньше, у них отсутствует защитный восковой щит и они никогда не размножаются. Гриб прикрепляется только к пораженному им насекомому, и его гифы никогда не проникают в субстрат. Рост гриба начинается весной (тогда образуется новая беловатая зона вокруг плодового тела) и продолжается до осени, а затем на поверхности новой зоны начинает развиваться гимений. В начале зимы образуются пробазидии, которые, пройдя период покоя, весной и летом следующего года, при достаточно влажной погоде дадут начало базидиям. Базидиоспоры отпадают от стеригм, и многие из них остаются на поверхности гриба, где они начинают почковаться.
Наиболее обильное спороношение наблюдается в мае. В это же время от перезимовавших самок рождаются личинки щитовок первого поколения. Одни личинки остаются жить под грибом, где они родились, другие передви-гаются в соседнюю колонию, а третьи выбирают себе место на коре, где нет гриба. Именно эти личинки и обеспечивают распространение гриба, так как, уходя из "дома", они переходят гимений и заражаются спорами, точнее, вторичными спорами, возникшими в результате почкования. Споры гриба приклеиваются к ногам личинки, прорастают, и его клетки проникают в полость тела насекомого.
В начальной стадии развития гриб существует в виде веретеновидных клеток, которые свободно плавают в гемолимфе личинки. Затем из них вырастают очень тонкие гифы. Они начинают расширяться до тех пор, пока вся полость тела личинки не заполняется цепями веретеновидных клеток, соединенных между собой тонкими нитями. Через две-три недели после первой линьки в теле насекомого начинают развиваться спиралевидные гаустории. Примерно в это же время гифы гриба прорастают через естественные отверстия тела насекомого наружу и после второй линьки сливаются, образуя над насекомыми покрывало. При этом соединяющие гифы имеют вид спиралей, так что гриб прикрепляется к щитовке как бы слабыми пружинками, что оставляет ей определенную свободу движения.
Через два месяца после заражения личинки колония гриба достигает диаметра 2 мм и толщины 1 мм. Для дальнейшего роста гриба необходимо, чтобы под ним нашли убежище новые личинки, зараженные спорами. Очевидно, гриб привлекает их запахом и благодаря позитивному хемотаксису щитовок обеспечивает себя необходимым источником питания. Пораженные насекомые живут дольше, чем непораженные. Таким образом распространение и рост септобазидиума зависят исключительно от щитовок, но совместная жизнь выгодна и насекомому. Здоровые щитовки никогда не заражаются гифами гриба и находят под ним защиту от невыгодных климатических условий, а также от своих врагов - птиц и перепончатокрылых насекомых-паразитов.
В европейской части СССР встречается септобазидиум карестианум (Septobasidium carestianum), который растет на ветвях ивы, ясеня, рябины и связан с европейской ивовой щитовкой (Chionaspis salicis). Плодовые тела этого гриба широкораспростертые, пленчатые, продолговатые, светло-коричневые, с беловатым краем. В течение вегетативного периода под плодовым телом гриба можно видеть зараженных и здоровых щитовок в различных стадиях развития. Так как из яиц, отложенных с осени, личинки появляются лишь весной, то вегетативный рост гриба зимой задерживается до весны, когда вылупившиеся личинки заражаются спорами и продолжают жить под колонией гриба. Насекомые, не зараженные спорами, уже никогда не будут заражены гифами гриба, под которыми они живут.
Многие субтропические и тропические виды рода септобазидиум считают фитопатогенными, так как ветви деревьев и кустарников, на которых они растут, часто высыхают. Это вызывается совместным действием насекомых и гриба. Химические меры борьбы остаются малоэффективными, потому что достаточно толстые плодовые тела остаются в нижней части жизнеспособ-ными и защищают щитовок от ядохимикатов, что еще раз доказывает взаимную зависимость жизни гриба и щитовок.
Род урединелла (Uredinella) содержит один вид U. coccidiphaga, паразитирующий на щитовках. Плодовые тела этого гриба - маленькие дисковидные пятна, диаметром до 1,5 мм. Одна колония этого гриба всегда живет только на одной особи насекомого. В отличие от представителей рода септобазидиум урединелла - однолетний организм. Осенью на поверхности колонии гриба образуются толстостенные бурые пробазидии, которые очень похожи на телейтоспоры ржавчинных грибов. Следующей весной из перезимовавших пробазидий или телейтоспор начинают развиваться четырехклеточные базидии, которые должны дать начало базидиоспорам, после чего старое плодовое тело погибает.
Телейтоспоры урединеллы всегда одноядерные. Рядом с ними встречаются похожие на них более продолговатые толстостенные бурые двухъядерные клетки, которые дадут начало крупным двухъядерным спорам. Эти клетки и споры рассматриваются как материнские клетки уредоспор и уредоспоры.
Семейство Аурикуляриевые (Aubiculabiaceae)
Плодовые тела представителей этого семейства имеют различную форму, от широкораспростертого или бугорковидного до булавовидного или уховидного. Базидия четырехклеточная, с поперечными перегородками. Споры одноклеточные, при прорастании образуют вторичные споры, мало отличающиеся от базидиоспор. Во многих родах встречаются сапротрофные и паразитные виды.
Представителем кортициевидной жизненной формы в этом семействе является род геликоглоя (Helicogloea). Грибы этого рода имеют широкораспростертые, сравнительно тонкие плодовые тела, причем в пределах рода наблюдается переход от сухой консистенции (Helicogloea farinaсеа) к студенистой (Н. lagerheimii). Оба гриба растут на нижней стороне поваленных гниющих стволов и очень похожи на кортициевые грибы.
Род платиглоя (Platygloea) объединяет грибы со студенистыми бугорковидными плодовыми телами. Многие виды являются сапротрофами, но встречаются и представители, паразитирующие на других грибах.
Платиглоя (Platygloea peniophorae) образует плоские дисковидные бугорки на плодовых телах многих кортициевых грибов. Растет он и на грибах рода дакримицес (Dacrymyces), но примечательно, что на их студенистых плодовых телах P. peniopliorae самостоятельных плодовых тел не образует. Базидии паразита в данном случае развиваются прямо в гимении хозяина. P. peniophorae - хорошо приспособленный паразит, он не вызывает деформации и некроза тканей гриба, на котором паразитирует.
Род эокронартиум (Eocronartium) представлен единственным видом Е. muscicola. Его тонкие булавовидные белые плодовые тела, высотой до 3 см и шириной до 3 мм, очень напоминают некоторых представителей рогатиковых грибов (рис. 210).
Е. muscicola развивается как паразит на живых мхах из родов климациум (Climacium), гипнум (Hypnum), амблистегиум (Amblystegium), туидиум (Thuidium) и др. Зачатки плодовых тел гриба можно обнаружить уже в начале лета в виде маленьких белых бугорков на верхушках зараженных стеблей мхов. При этом зараженные мхи ничем не отличаются от здоровых и способны продолжать свой рост. Однако уже при поверхностном осмотре видно, что плодовые тела гриба берут начало от мицелия, который развивается внутри растения-хозяина. При микроскопическом анализе выявляется, что стебель и ветви зараженного мха заполнены мицелием гриба. Гифы Е. muscicola развиваются в тканях мха внутриклеточно, и их можно обнаружить практически в каждой клетке, но никаких гаусториев они не образуют. Зараженные клетки содержат живую цитоплазму и вполне развитое ядро. Если сравнить их с клетками незараженного мха, то трудно заметить какие-либо существенные отклонения. Все это свидетельствует о том, что Е. muscicola является очень хорошо приспособленным паразитом.
Мицелий гриба многолетний. Каждой весной в зараженных мхах можно найти как старые, прошлогодние, так и молодые гифы, которые проникают в молодые побеги мха и летом образуют плодовые тела на их вершинах. Перед образованием плодового тела гифы вырастают из верхней части побега и образуют над листьями плоские мицелиальные пленки. Эти пленки удлиняются, становятся похожими на волокна и, наконец, покрывают верхушку побега. Затем они срастаются, образуя ножку плодового тела, на вершине которого появляется фертильная головка. На поверхности головки развиваются типичные для порядка четырехклеточные базидии с поперечными перегородками. Период споруляции длится от двух до трех недель.
Род геликобазидиум (Iielicobasidium) объединяет грибы, для которых характерны широко распростертые плодовые тела и сильно искривленные базидии.
Геликобазидиум пурпурный (H. purpureum) имеет мясисто-волокнистое с фиолетовым или пурпурным гимением плодовое тело, диаметром 5 см. Растет гриб как сапрофит на гниющей древесине, но несовершенная стадия этого вида (Rhizoctonia crocorum) паразитирует на корнях многих кормовых и овощных культур, например свеклы, моркови, люцерны.
Все грибы рода аурикулярия (Auricularia) имеют крупные уховидные студенистые плодовые тела и все являются сапрофитами на мертвой древесине. Однако есть сведения, что самый распространенный вид в европейской части СССР и в Западной Европе A. mesenteric может расти и как факультативный паразит, иногда сильно поражая ослабленные фруктовые деревья и вызывая их гибель. Таким образом, во всех родах семейства аурикуляриевых видны тенденции к паразитическому образу жизни.
A. mesenterica примечательна еще и тем, что хотя она и может расти на гниющей древесине многих лиственных пород, однако чаще поселяется на древесине вязов (Ulmus), особенно на их поваленных стволах.
Аурикулярией богат Дальний Восток. Туда проникают ареалы таких субтропических видов Юго-Восточной Азии и Средней Америки, как A. cornea и A. polytricha. В Японии эти грибы считаются излюбленными съедобными грибами и известны под названием "древесные уши".
Семейство Флеогеновые (Phleogenaceae)
В это семейство входит лишь один род флеогена (Phleogena) с единственным видом Phleogena faginea, который выделяется среди всех представителей порядка аурикуляриевых замкнутыми ангиокарпными плодовыми телами, подобными плодовым телам гастеромицетов.
Phleogena faginea (рис. 209) встречается на отмершей коре лиственных и очень редко хвойных деревьев. Его плодовые тела имеют ножку высотой 5-7 мм, которая заканчивается шаровидной головкой диаметром до 1-3 мм. Головка покрыта корковидным перидием, и внутри ее развиваются базидии и споры. В отличие от других аурикуляриевых грибов на базидиях P. faginea отсутствуют стеригмы, так же как и у некоторых гастеромицетов. Толстостенные шаровидные желтовато-бурые споры образуются непосредственно на клетках базидия. Таким образом, споры P. faginea более похожи на споры гастеромицетов, чем на споры других аурикуляриевых грибов.
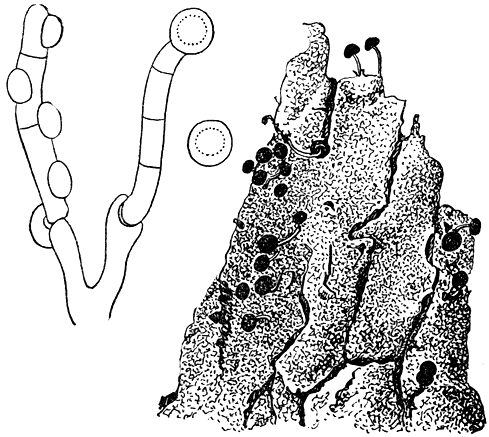
Рис. 209. Флеогена фагинея (Phleogena faginea): справа - плодовые тела; слева - базидии и споры
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'