Порядок Сферейные (Sphaeriales) (К. Л. Тарасов)
Сферейные - один из наиболее обширных порядков среди грибов, насчитывающий несколько тысяч видов. Относящиеся сюда организмы чрезвычайно широко распространены в природе и часто встречаются в огромном количестве в самых различных местообитаниях. Преимущественно это сапрофиты, хотя встречаются и паразиты, иногда даже очень опасные. Однако большинство из этих столь нередких грибов малозаметны. Ботаник, не занимающийся специально микологией, может даже не обратить внимания на черные или темно-коричневые точки, столь обычные на сухой траве, древесине и т. д. А между тем очень часто именно так выглядят плодовые тела или более крупные мицелиальные образования - стромы - у этих грибов. На почве же или навозе, где еще чаще встречаются представители данного порядка, они вообще незаметны.
Плодовые тела здесь почти всегда - типичные перитеции. Это мелкие (как правило, диаметром не более 2 мм), округлые или грушевидные тельца с полостью внутри, сообщающейся с внешним миром при помощи узкого отверстия на вершине, иногда оттянутой в носик. Отверстие изнутри выложено мелкими волосковидными выростами - перифизами. Стенки перитециев многослойные. Перитеции могут быть свободными и расположен-ными под тканью растения, на котором сапрофитно или паразитно обитает гриб. Также они могут быть погруженными в специальные тканевые образования - стромы. На дне полости перитеция располагаются булавовидные или цилиндрические сумки, иногда в виде пучка, иногда плотным слоем; большей частью они перемешаны с бесцветными парафизами. Сумки, постепенно созревая, вытягиваются, внедряются верхушкой в отверстие перитеция и выбрасывают наружу свои споры. После этого оболочка сумки спадает и ее место занимает следующая. У более примитивных представителей активного выбрасывания спор из перитеция не происходит и споры вследствие расплывания оболочек сумок оказываются прямо в его полости. В дальнейшем они выходят наружу через отверстие перитеция в общей взбухающей слизи или освобождаются при разрушении его стенки.
Для большинства сферейных характерно наличие в сумке специальной поры, предназначенной для активного удаления спор. Ее устройство иногда бывает очень сложным, напри- мер у грибов семейства ксиляриевых (Xylariaceae) - так называемый апикальный аппарат.
У сферейных оболочка перитеция и строма (где она имеется), за редким исключением, черные или темных тонов. Им свойственна углистая консистенция.
Половой процесс может быть различным. Здесь мы встречаем как примеры типичной для сумчатых грибов картины, так и всевозможные отклонения. Антеридий, например, часто не функционирует или вообще утрачен. Встречается также сперматизация - оплодотворение аскогона самостоятельными клетками - спермациями. Последние чаще образуются экзогенно, по типу конидий, но у некоторых видов возникают эндогенно.
Многие сферейные не имеют конидиального спороношения, но у других оно хорошо развито и может даже преобладать в цикле развития. У некоторых представителей (особенно у паразитических видов) хорошо выражен плеоморфизм - обилие различных конидиальных форм у одного и того же организма.
Перитеции, как и у многих других сумчатых грибов, приходят обычно на смену конидиальному спороношению, характерному в умеренном климате для летнего времени. У паразитов перитеции часто развиваются уже тогда, когда ткань растения-хозяина убита, и гриб перешел к сапрофитному образу жизни.
В настоящее время в порядке сферейных известно около 18 семейств. Некоторые из них мы рассмотрим в ходе дальнейшего изложения. Наиболее примитивные еще мало отличаются от эуроциевых: отсутствует активное выбрасывание спор из сумок, парафиз нет или они быстро исчезают. Встречаются грибы без выходного отверстия перитеция. Таковы семейства хетомиевых (Chaetomiaceae) и меланоспоровых (Melanosporaceae). От хетомиевых берет начало центральное семейство порядка - сордариевые (Sordariaceae). Здесь мы имеем типичные перитеции, свободные или погруженные в субстрат. Строма здесь отсутствует, но сумки имеют вершинные утолщения, связанные с активным выбрасыванием спор. От сордариевых идут две ветви сферейных, имеющих строму. Одна ветвь представлена семействами розеллиниевых (Kosellinmceae) и ксиляриевых (Xylariaceae), у представителей которых строма составлена исключительно из гиф гриба. Апикальный аппарат достигает очень высокого развития. Другая ветвь, идущая от сордариевых, характеризуется тем, что строма образуется не только из гиф гриба, но также из тканей растения, на котором гриб паразитирует или существует сапрофитно. Таковы семейства филлахоровых (Phyllachoraceae), полистигмовых (Polystigmataceae) и диатриповых (Diatryрасеае).
Семейство Хетомиевые (Chaetomiaceae)
К семейству хетомиевых (Chaetomiaceae) относятся наиболее примитивные представители порядка. Эти грибы весьма близки к представителям порядка эуроциевых, поэтому некоторые исследователи вообще относят сюда хетомиевых. Некоторые хетомиевые еще не имеют выходного отверстия перитеция. В перитеции отсутствуют парафизы. Оболочка сумок по всей поверхности равной толщины, без специальных образований, связанных с освобождением спор. Она рано разрушается, и зрелый перитеций наполняется массой аскоспор, которые затем в общей слизи выступают наружу. Подобные особенности заставляют считать хетомиевые переходной группой от эуроциевых к типичным сферейным.
Хетомиевые - сапрофиты, обитающие преимущественно на растительных остатках. Они играют определенную роль в разрушении растительного опада на почве (некоторые из них разрушают материалы), обитают на навозе.
На различных субстратах нередко встречаются виды обширного рода хетомиум (Chaetomium). Они характеризуются наличием у перитециев волосков, обычно закрученных, простых или разветвленных (рис. 92). Последние могут служить хорошим диагностическим признаком при идентификации видов. Например, они могут быть волнистыми по всей длине или, как у С. murorum, делать на вершине только один или два спиральных завитка; могут оканчиваться бесцветными вздутиями, как у С. zopfii. Окраска их тоже различна: у С. murorum, например, она зеленоватая, у С. cochlioides темно-лимонная, а у одного из его мутантов розовая.
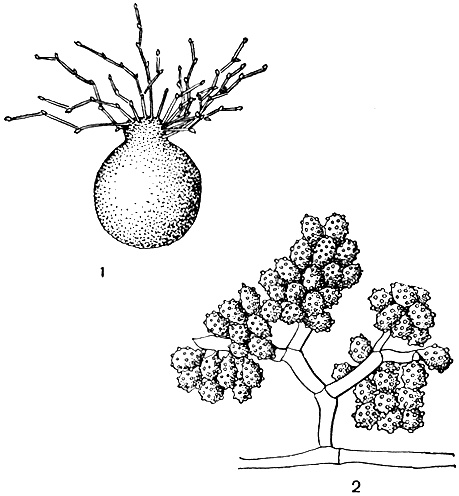
Рис. 92. Хетомиум (Chaetomium): 1 - внешний вид перитеция; 2 - конидиальное спороношение
Некоторые виды (например, С. globosum) наносят определенный ущерб хозяйству, разрушая клееные конструкции (фанеру), древесину, бумагу, особенно в условиях жаркого и влажного климата. Среди грибов, являющихся причиной заплесневения одежды, также есть представители этого рода.
Конидиальные спороношения в этом семействе встречаются далеко не у всех видов и могут быть самыми различными.
Семейство Меланоспоровые (Melanоsporaceae)
Второе, тоже довольно примитивное семейство этого порядка - меланоспоровые (Melanosporaceae). Сумки у его представителей, как и у хетомиевых, рано ослизняются. Также ослизняются уже имеющиеся здесь парафизы.
Грибы рода меланоспора (Melanospora) - сапрофиты на всевозможных разлагающихся субстратах животного и растительного происхождения, реже - паразиты на насекомых или грибах, в свою очередь также паразитирующих на насекомых. Перитеции свободные или погруженные в сплетение гиф. Оболочка зрелых перитециев однослойная, коричневатая или даже светлая, мягкая. Вершина их обычно оттянута в длинный носик (рис. 93) с отверстием на вершине, через которое выталкивается масса аскоспор, освобождающихся при разрушении сумок. Споры коричневатые, с характерным рельефным сетевидным рисунком. Имеется проростковая пора. Клетки гиф многоядерные.
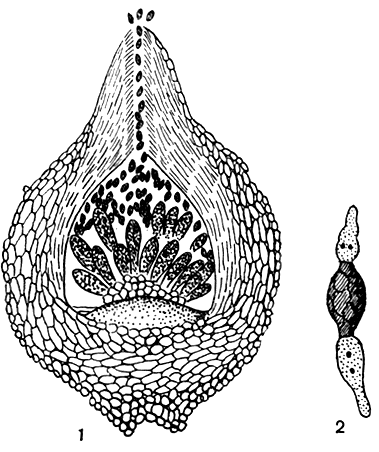
Рис. 93. Меланоспора (Melanospora): 1 - перитеций; 2 - прорастающая аскоспора
Семейство Сордариевые (Sordariaceae)
Типичные представители сферейных - формы, объединяемые в семейство сордариевых (Sordariaceae). Разрушения стенок сумок здесь не происходит и споры активно разбрасываются. В большинстве своем это широко распространенные копрофильные грибы, развивающиеся на навозе травоядных животных. Перитеции их черные или темно-коричневые, высотой 1-2 мм. Они появляются обычно на смену мукоровым (Mucorales). Перитеции свободные или погруженные в субстрат, гладкие или покрытые мягкими волосками. Цилиндрические сумки внутри них перемежаются с многочисленными парафизами. Однако к моменту созревания спор у многих грибов парафизы исчезают. Споры коричнево-черные, эллиптические, одноклеточные, часто со слизистыми придатками. Характерно наличие специального кольца или круглого валика на вершине сумки - приспособления для активного выбрасывания спор.
С экономической точки зрения это семейство не имеет большого значения, однако некоторые грибы служат излюбленными объектами лабораторных опытов.
Представители рода сордария (Sordaria) - копрофильные грибы. Они хорошо растут в культуре на искусственных субстратах. Верхняя часть их сумок снабжена вполне развитым кольцом. Каждая спора имеет одну проростковую пору. Остальная часть споры со всех сторон окружена слоем воднорастворимой слизи.
S. fimicola не имеет конидиального спороношения и размножается только аскоспорами. Ученые проводят детальные исследования в области физиологии и генетики этого гомоталличного вида.
Гриб этот образует перитеции рано и в большом количестве, если содержание углеводов становится неблагоприятным для вегетативного роста. Образование плодовых тел обильнее при рН больше 6. Образованию перитециев способствует биотин, у некоторых штаммов - тиамин. Биотин служит существенным фактором не только для образования перитециев, но и для образования и созревания сумок. При очень низких концентрациях биотина возникают перитеции без сумок. Наблюдается прямая взаимосвязь между количеством добавляемого биотина и процентом образующихся зрелых аскоспор.
Шейки перитециев у сордарии, как у большинства представителей семейства, положительно фототропичны.
К этому роду очень близки роды гелазиноспора (Gelasinospora) и нейроспора (Neurospora).
Грибы рода гелазиноспора, как и другие представители семейства,- быстрорастущие грибы. Клетки мицелия у них многоядерные, причем ядра могут переходить через поперечные перегородки из клетки в клетку довольно легко. Мицелий окрашенный, количество пигмента зависит от вида субстрата. Как и многие другие грибы, виды этого рода нуждаются в биотине для роста и размножения. Ни один из известных представителей рода гелазиноспора не образует настоящих конидий. У некоторых бывают микроконидии, но их роль в воспроизведении вида сомнительна: в культуре они прорастают плохо и растут медленно.
Проводились опыты по скрещиванию Gelasinospora autosteria с Sordaria brevicaulis. Оба вида гетероталличны. В местах соприкосновения мицелиев в некоторых случаях в изобилии возникали перитеции, но сумки в них не развивались.
Из представителей этого рода можно назвать G. cerealis - почвенный гриб, возможно, вызывающий полегание пшеницы и овса. Его аскоспоры (в сумке их 8, как и у большинства сумчатых грибов) к моменту созревания становятся двуклеточными. У копрофильного вида G. tetrasperma каждая сумка имеет по 4 споры, в каждой из которых содержится по 2 ядра.
Грибы рода нейроспора чаще встречаются в конидиальной стадии, образуя микроконидии, а также скопления оранжевых или розовых конидий. Мицелий состоит из обильно ветвящихся гиф, клетки которых всегда одно-ядерные. Перитеции здесь, в отличие от большинства сферейных, светлые. Эти грибы широко распространены в природе (особенно в почве) и издавна являются излюбленными объектами генетических исследований. Такой вид, как нейроспора густая (N. crassa), - пожалуй, наиболее популярный объект среди гаплоидных организмов, подобно тому как муха дрозофила - среди диплоидных.
На нейроспоре густой был особенно успешно применен тетрадный анализ, т. е. анализ, проводимый по гаплоидным продуктам мейоза. Этот метод дает возможность анализировать гаплоидные особи, развивающиеся из аскоспор; он впервые позволил непосредственно доказать, что менделевское расщепление является закономерным ходом мейоза, что оно представляет не статистическую, а биологическую закономерность. Этот метод позволил определить результаты кроссинговера непосредственно по гаплоидным продуктам мейоза, что необходимо для доказательства соответствия рекомбинантных зигот кроссинговерным гаметам.
Гриб N. sitophila издавна в изобилии встречался в пекарнях. На рынках Явы долгое время пользовались спросом арахисные пирожные, окрашенные в оранжевый цвет этим грибом. Он также использовался в лабораториях, но очень скоро стал настоящим бичом, так как благодаря легко разносимым конидиям и быстрому росту он моментально распространялся по помещениям, заражая культуры (подобно злостным сорнякам на полях) и срывая опыты. Поскольку этот гриб нередко выделяется из почвы, все время существует опасность появления его в лабораториях, где так или иначе имеют дело с почвой. В случае его появления принимают самые строгие меры: временное прекращение работ, обработку помещения и посуды формалином и т. д.
Перитеции нейроспоры часто бывают недоразвитыми. В этом случае их называют склероциями.
У сордариевых, группируемых в род плевраге (Pleurage), споры снабжены шнуровидными слизистыми придатками, способствующими приклеиванию их к окружающим предметам, главным образом к траве.
Семейство Розеллиниевые (Roselliniaceae)
Наиболее примитивные представители строматических сферейных объединяются в семейство розеллиниевых. Строма у них развита еще очень слабо. Некоторые микологи даже не считают за строму рыхлое сплетение гиф, характерное для этих грибов.
Из разных частей света известно около 200 видов рода розеллиния (Rosellinia), встречающихся преимущественно на древесине и коре (рис. 94). Подавляющее большинство этих грибов - сапрофиты, но есть и несколько опасных паразитов. Например, R. aquila вызывает загнивание шелковицы, R. quercina поражает дуб, R. necatrix заражает виноградные лозы, а также плодовые деревья (яблони, груши, сливы, абрикосы), иногда шелковицу и кофейное дерево. После того как пораженные органы растения-хозяина (преимущественно корни) отмирают, гриб продолжает свое развитие, образуя коричневые склероции, превращающиеся затем в пикниды с конидиеносцами, отчленяющими бесцветные конидии. Наконец, образуются перитеции, почти округлые, только на вершине оттянутые в сосочковидный выступ, на котором имеется отверстие. Основание их более или менее погружено в рыхлое сплетение гиф. Между сумками расположены многочисленные нитевидные парафизы.
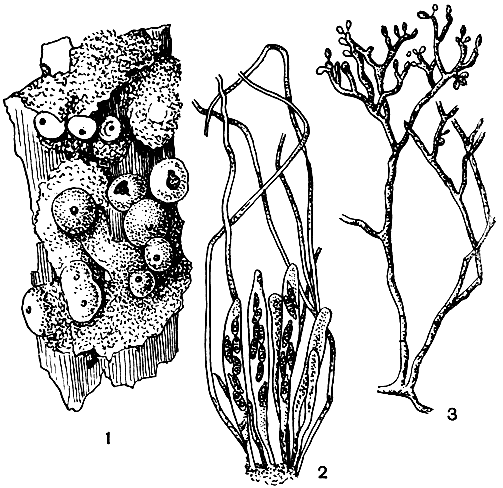
Рис. 94. Розеллиния (Rosellinia): 1 - перитепий; 2 - сумки и парафизы; 3 - конидиальное спороношение
От R. quercina страдают молодые дубы. Гриб поражает их корни, почему и называется "дубовым корнегубителем". Болезнь эта наиболее распространяется на северо-западе ФРГ в сырые, дождливые годы. Молодые (однолетние и трехлетние) дубки увядают, сохнут и гибнут. Отмирание начинается с верхушки стеблей. На корнях увядающего растения можно разглядеть черные шарики величиной с булавочную головку - склероции. Кроме того, местами на корне заметны еще нежные, нитевидные, ветвящиеся тяжи, состоящие из слабо сросшихся гиф. Они протягиваются в почве от одного корня к другому и служат для распространения гриба. Если дубки со склероциями посажены летом в сырую почву, то склероции дают начало паутинистому мицелию, который распространяется по поверхности почвы. Гифы, достигнув корней, обвивают их, проникают внутрь до самой сердцевины и довольно быстро убивают дерево. В живых паренхимных клетках коры гифы разрастаются, заполняя их полость плотным сплетением. Старые части корня защищены от проникновения мицелия пробковой тканью, но в случае повреждения (особенно при отрывании боковых корешков) они могут также поражаться. Летом на внешнем мицелии, образовавшемся из склероция, развиваются конидиеносцы с конидиями. На таком же мицелии развиваются перитеции в виде черных точек. Аскоспоры прорастают только на следующий год, тогда как конидии прорастают тем же летом и обычно разносятся мышами.
Достаточно распространена во многих странах Старого и Нового Света так называемая "плесневая болезнь", или "белая гниль", винограда и фруктовых деревьев, вызываемая R. necatrix. Она известна во Франции, Алжире, Тунисе, США (штаты Миссури, Пенсильвания, Северная и Южная Каролина, Техас, Калифорния), а также во многих других местах.
В нашей стране встречается в первую очередь в Крыму и Молдавии.
Все разводимые виды и сорта винограда подвержены этому заболеванию (конечно, в разной степени). Этим грибом также поражаются почти все плодовые деревья, помимо этого круг растений-хозяев гриба охватывает розы, дуб, бобы, картофель, ель, сосну и другие растения. Наиболее сильно гриб развивается в сырых местах, чаще всего на глинистых и мергелистых почвах, переувлажненных. На таких почвах высшие растения бывают ослаблены из-за недостатка железа и марганца. Хотя общее количество железа может быть велико (так же как и марганца), эти элементы, особенно железо, находятся в форме двухвалентных ионов и не только недоступны для растения, но и оказывают токсическое действие. По склонам холмов, на аллювиальных песках, известковых и гранитных почвах заболевание растений наблюдается сравнительно реже.
Первоначально поражение винограда этим грибом легко можно спутать с поражением филлоксерой или с хлорозом. Заболевшее растение у основания начинает более обильно ветвиться. Листья его становятся мельче, более изрезанными, а потом желтеют и отмирают. В дальнейшем основание стебля становится бурым, ноздреватым и с этого места легко отпадает кора. Если срезать стебель у основания, из среза в изобилии, особенно весной и осенью, вытекает черное вещество, дающее реакции на сахар. Виноград погибает через 15-19 месяцев, фруктовые деревья - через 24-36 месяцев. Это заболевание передается от одного растения к другому сравнительно медленно, но в густых насаждениях оно может распространяться быстрее.
Гриб может жить в почве как сапрофит. При благоприятных условиях мицелий появляется на поверхности корней, а также в виде тяжей, похожих на корни,- ризоморф (подобно ризоморфам опенка). Достигнув молодого корня, гриб убивает его и затем проникает в более старый корень, разрастаясь в его коре также в виде ризоморф, теперь уже внутренних. Ризоморфа ветвится, образуя клубневидные склероции и разрывая местами кору, где выходит наружу и развивается либо в паутинистый мицелий, либо в ризоморфу. Во время паразитной жизни гриба никаких спороношений не развивается. Только после гибели растения-хозяина, в сапрофитных условиях, он начинает развивать конидиеносцы с конидиями и плодовые тела, покрывая ими поверхность пораженных органов.
Ветвистые темные конидиеносцы появляются пучками на склероциях и мицелии (рис. 94,3). Перитеции появляются по нескольку на склероциях. Они почти шаровидные, сверху слегка сплюснутые, с небольшим носиком. Оболочка их черная, углистая, ломкая. Споры черно-бурые, одноклеточные. Сумки перемежаются с многочисленными парафизами (рис. 94,1,2).
В качестве мер борьбы может быть эффективным дренирование почвы, а также уничтожение больных растений до их гибели, пока гриб не начал развивать спороношений.
Семейство Ксиляриевые (Xylariaceae)
Наиболее развитые стромы, состоящие исключительно из гиф, характерны для грибов из обширного семейства ксиляриевых (Xylariaсеае).
У грибов рода нуммулярия (Nummularia) строма округлая, дисковидная, аморфно-корковатая, черная. Таков гриб нуммулярия дискретная (N. discreta). Он обычно встречается на засохших ветвях и стволах рябины, яблони и других лиственных пород, где образует стромы в виде выступающей из-под коры плоской, черной, округлой чашечки с утолщенным шероховатым краем. Этот гриб не всегда существует сапрофитно и иногда поражает живые яблони, вызывая у них так называемый "иллинойский рак". Для стран Старого Света это заболевание не указывалось.
У других грибов строма развивается целиком на поверхности. Однако у устулины (Ustulina) она, как и у представителей предыдущего рода, еще не имеет определенной формы. U. vulgaris покрывает поверхность стволов и ветвей сухих деревьев черными волнистыми корками часто внушительных размеров. Сначала они мягкие и покрыты конидиальным гимением, но затем становятся хрупкими, черными, углистыми. Близкий вид U. zonata вызывает заболевание корней гевеи и чайных кустов.
У более развитых родов, таких, как гипоксилон (Hypoxylon) (рис. 95, табл. 18) и далъдиния (Daldinia), стромы постепенно приобретают более четкую форму. Они полукруглые или клубневидные. Однако молодые стромы выглядят еще в виде корковидных образований, напоминая стромы устулины. В некоторых случаях они могут оставаться такими. На мицелии обоих этих родов образуются очень изящные, часто характерно ветвящиеся конидиеносцы. Конидии возникают или по бокам их, как у Hypoxylon unitum, или, как у Н. fuscum и Н. coccineum, на вершине, в плотной небольшой головке. Иногда конидиеносцы группируются в коремии. Обычно конидиеносцы встречаются по всему мицелию; реже они приурочены к поверхности молодой стромы. Во всех случаях перитеции развиваются после исчезновения конидиального спороношения.
Оба вида обитают в основном на гниющей древесине и сухих ветвях. У гипоксилона стромы гомогенные, у дальдинии они состоят из концентрических кругов. Отсюда и название очень обычного представителя этого рода - дальдиния концентрическая (D. concentrica).
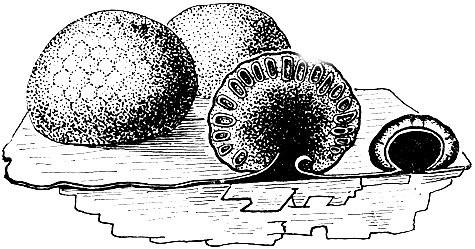
Рис. 95. Гипоксилон (Hypoxylon). Внешний вид стромы
Наиболее развитыми в этом семействе являются грибы родов ксилярия (Xylaria), тамномицес (Thamnomyces) и порония (Poronia). Строма их расчленена на стерильную и фертильную части.
Род ксилярия - космополит, но особенно широко представлен в тропиках. Грибы этого рода встречаются преимущественно на мертвой древесине, значительно реже на навозе или сухих плодах (например, X. carpophila - на орешках бука). Из аскоспор развивается обширный мицелий, гифы которого объединяются в толстые тяжи. Последние показывают интенсивный гелиотропизм, так что, даже начав развиваться под корой или стволом дерева, они легко выходят на открытое пространство. Постепенно они развиваются в цилиндрические булавовидные или ветвящиеся стромы (рис. 96, табл. 18). Растущая вершина стромы долгое время остается белой, покрытой правильным палисадным слоем конидиеносцев. Последние бывают одноклеточными и многоклеточными. В первом случае они отчленяют яйцевидные конидии, во втором на их вершине образуются веретеновидные конидии. У грибов двух американских видов (X. tentaculata и X. tracheiina) конидиальные структуры возникают не непосредственно от стромы, а на специальных веточках, которые растут наподобие коремий от разветвлений стромы. Спустя большой промежуток времени, после того как исчезнут конидиальные структуры (обычно весной следующего года), ветви стромы булавовидно утолщаются в верхней части и образуют перитеции.
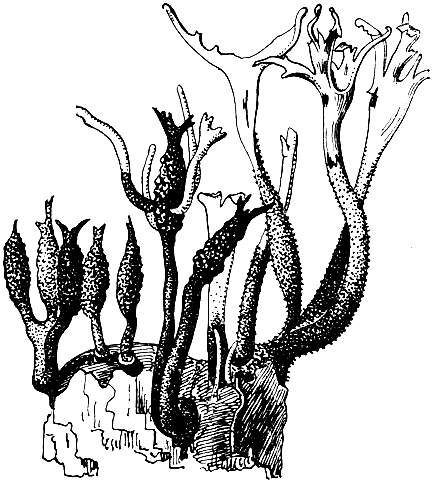
Рис. 96. Ксилярия (Xylaria). Внешний вид стромы

Таблица 18. Сферейные и диапортовые грибы: 1 - лист грецкого ореха, пораженный гномонией тонкозаостренной (Gnomonia leptostyla): вверху слева - конидиальное спороношение (ложе); вверху справа - перитеции; 2 - поражение плода грецкого ореха тем же грибом; 3, 4 - разветвленные стромы различных видов ксилярии (Xylaria); 5 - гипоксилон (Hypoxylon)
Thamnomyces chamissonis (рис. 97) развивается на сухой, еще не разложившейся древесине, лежащей на земле в лесах. Стромы его растут сперва в виде тонких (диаметром 1-2 мм), волосовидных образований, группирующихся в кустики, иногда довольно большие. Примерно до высоты 7 см они растут не ветвясь. Затем они последовательно 5-6 раз дихотомически ветвятся, причем ветвь каждого последующего порядка тоньше и короче предшествующей. Таким образом возникают "деревца" до 11 см высоты. Каждая из конечных веточек слегка утолщается и содержит по одному перитецию с плотной углистой стенкой.
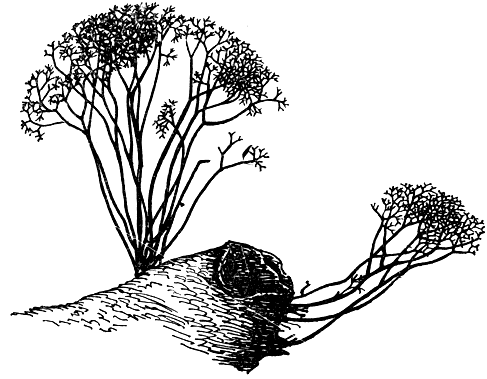
Рис. 97. Тамномицес (Thamnomyces). Внешний вид стромы
Род порония отличается от других родов семейства: у его грибов спороносная часть стромы дисковидно разрастается. Наиболее распространенный в северном полушарии вид этого рода - P. punctata - встречается на старом конском навозе, из которого часто высовываются только фертильные диски гриба. Сперва на них развиваются светло-серые конидии, позже - перитеции.
Среди семейств, где строма составлена не только из гиф гриба, но и с привлечением тканей растения-хозяина, также можно выделить более примитивные и более высокоорганизованные.
Семейство Филлахоровые (Рнyllachoraceae)
Переходной группой от сферейных со свободными перитециями к формам с перитециями, погруженными в строму, в данном случае является семейство филлахоровых (Phyllachoraсеае). Некоторые грибы единственного относимого сюда рода филлахора (Phyllachora) при определенных условиях образуют строму.
Черные округлые перитеции погружены в ткань листа. Стенки сумок равномерно утолщены и имеют по большой поре на вершине. Сумки перемежаются с многочисленными нитевидными парафизами.
В роде около 200 видов. Из них широко известна злаковая филлахора (P. graminis), паразитирующая на злаках. Этот гриб образует на листьях многих злаков мелкие, слегка блестящие роговые пятна (рис. 98). Он встречается преимущественно на пырее, просе, еже, костре, тимофеевке, а также на осоке и ожике. Обычно все листья пораженного растения бывают покрыты пятнами, являющимися стромами разнообразной величины и формы. Стромы состоят из многочисленных, очень тонких гиф, идущих между клетками листовой паренхимы. Сосудистые пучки при этом остаются нетронутыми. Гифы заполняют внутри и клетки эпидермиса, образуя прочный покровны"й слой.
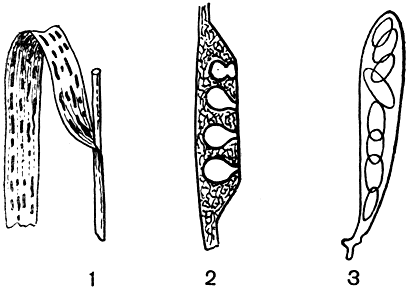
Рис. 98. Филлахора злаковая (Phyllachora graminis): 1 - пораженное растение; 2 - разрез стромы с перитециями; 3 - сумка
В сырую погоду, особенно во влажных местах, значительный вред клеверу может причинить P. trifolii. Гриб образует на нижней стороне листьев многочисленные мелкие черные матовые пятна, вследствие чего болезнь известна как "почернение клевера". Каждое пятно состоит из многочисленных полушаровидных бугорков, представляющих собой пучок прямостоячих конидиеносцев, выходящих через разрыв эпидермиса листа. Некоторое время пораженные листья остаются зелеными, потом желтеют и сохнут. К осени, во время листопада, на них под конидиальной стромой образуется другая, черная строма. На ней развиваются сначала пикниды, потом перитеции.
Семейство Полистигмовые (Polystigmatacеае)
К филлахоровым близки представители семейства полистигмовых (Polystigmataceae). Перитеции и стромы их также располагаются внутри растения-хозяина, только строма более развита.
На листьях сливы, вишни, черешни можно наблюдать ярко-красные мясистые пятна, видимые с обеих сторон листа. Иногда эти пятна мелкие, иногда занимают половину листа или целый лист. Это стромы гриба полистигма (Polystigma). Летом в них развиваются пикниды с изогнутыми конидиями, осенью и весной, уже на опавших листьях,- перитеции. Аскоспоры бесцветные, эллиптические. Попав на молодые листья, они прорастают, вызывая новое поражение.
Наиболее распространен вид P. rubrum - возбудитель "ожога" листьев черешни, сливы и вишни. Мицелий, разрастаясь в ткани листа, гипертрофирует ее: мезофилл сильно разрастается и не содержит хлорофилла. Между его клетками и через них проходят красные ветвистые гифы, которые придают красный цвет пятну. Летом на нижней стороне пораженной части листа бывают заметны многочисленные точечные отверстия, ведущие в грушевидные полости,- пикниды, по стенкам которых отчленяются многочисленные изогнутые конидии. Они выходят на поверхность в виде беловатой капельки. В июле - августе в строме закладываются спиральные аскогоны, лишенные трихогины. Антеридии отсутствуют. Копуляция осуществляется приблизительно в декабре на отмерших листьях на земле. При этом между двумя сестринскими клетками аскогона (между многоядерной "мужской" и одноядерной "женской") возникает пора, через которую одно из "мужских" ядер переходит в "женскую" клетку и образует с ее ядром дикарион. Дальнейшее развитие аскогенных гиф и перитеция идет нормальным путем. Строма при этом буреет. Весной перитеции уже зрелые, и из них начинают выбрасываться эллиптические бесцветные аскоспоры. Попав на молодой лист, спора прорастает, и уже через 6 недель на этом месте возникает пятно с пикнидами.
Конидии (пикноспоры), развивающиеся в пикнидах P. rubrum, прорастают с большим трудом. Предполагают, что они являются спермациями, утратившими в настоящее время свою функцию и неспособными оплодотворять аскогон.
В случае массового развития гриб может причинять растениям существенный вред. Некоторую пользу для предупреждения заболевания может оказать сбор и сжигание опавших пораженных листьев.
Семейство Диатриповые (Diatrypaceae)
Одним из характерных признаков семейства диатриповых является то, что аскоспоры этих грибов часто имеют вид колбасы с перетяжками (аллантоидные). Обычно они одноклеточные, желтоватого цвета. Сумки булавовидные или цилиндрические, с длинными, суживающимися основаниями. В перитеции они образуют отчетливый гимениальный слой, где перемежаются с парафизами. Последние со временем ослизняются.
У наиболее примитивных форм этого семейства - грибов рода эутипа (Eutypa), обитающих на лишенной коры древесине, перитеции еще практически свободны. Их объединяет только черная корка мицелиального происхождения.
Главный род семейства - диатрипе (Diatryре), насчитывающий свыше 100 видов. У этих грибов довольно четко выражено разделение стромы на верхнюю и нижнюю части. Верхняя, сосочкообразная часть (эктострома) залагается под перидермой дерева и служит главным образом для ее прорыва. Кругом основания эктостромы, на плоском ложе, образуются конидии. Позднее, после ее разрушения или отпадения, разрастается закладывающаяся под ней внутренняя часть стромы (эндострома), в которой развиваются перитеции (рис. 99).
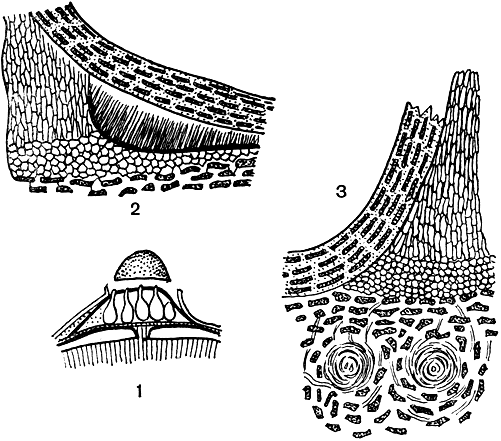
Рис. 99. Диатрипе (Diatrype): 1 - схематический разрез через экто- и эндострому; 2 - разрез части эктостромы с конидиальным спороношением у ее основания; 3 - разрез развивающейся эндостромы
D. disciformis обитает в качестве паразита на ветвях бука, впоследствии развиваясь уже сапрофитно на убитых тканях. Строма гриба плоская, диаметром около 2 мм, располагается она между паренхимой коры и перидермой хозяина. Возникающая сначала эктострома несет на себе конидиальное спороношение. Вследствие своей твердой консистенции и быстрого роста она оказывает такое давление на перидерму, что та приподнимается в виде плоских пустул. По краю стромы расположены полости, внутрь которых растут конидиеносцы, отделяя огромное количество мелких гиалиновых конидий, в массе кажущихся желтыми или мясисто-красными. За счет растяжения перидерма лопается и конидии высвобождаются через разрыв. Постепенно эктострома расплывается и отмирает, и в то же время развивается эндострома, в которой залагаются перитеции (их около 50). Верхний слой эндостромы превращается в жесткую корку, окружающую шейки перитециев.
У грибов, относящихся к родам криптосферия (Cryptosphaeria), валъсария (Valsaria) и антостома (Anthostoma), наблюдается тенденция к редукции эндостромы. Ножки сумок становятся короче, при этом увеличиваются в размере нитевидные парафизы.
Очень часто у строматических сферейных развитие стром наблюдается только при благоприятных условиях. В случае внезапного наступления засухи развитие может остановиться на какой-либо из начальных стадий, так что наблюдатель может обнаружить только стерильную склероциальную плектенхиму под перридермой, иногда с остатками конидиального спороношения. При этом различия между экто- и эндостромой практически стираются.
Многие представители этого порядка перешли к совместному существованию с водорослями и в настоящее время уже не существуют самостоятельно. Они составляют основную массу пиреномицетных лишайников (Pyrenolichenes) и будут упомянуты при рассмотрении лишайников.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'