Глава 5. Индукция и биосинтез фитоалексинов
Мысль о том, что после инфекции растения выделяют защитные химические вещества, высказывалась рядом исследователей еще в первой половине этого столетия, однако четко эта идея была сформулирована Мюллером и Бергером [179]. Как было показано в главе 4, к выводу о существовании фитоалексинов авторов привело изучение реакции сверхчувствительности у сортов картофеля, устойчивых к картофельной гнили, и эксперименты по индуцированию устойчивости у восприимчивых сортов. Термин "фитоалексин" был введен для означения образуемых растениями защитных соединений. Кроме того, считалось, что фитоалексин образуется в сверхчувствительных клетках тканей картофеля и мешает дальнейшему росту инфекционных гиф, причем предполагалось, что фитоалексины могут образовываться только в инфицированных растениях.
Мюллер [177], изучая реакции сверхчувствительности тканей боба к патогену мягких плодов Monilinia fructicola, впервые предпринял попытку выяснить химическую природу фитоалексина. Капельки суспензии спор помещали в полости открытых створок бобов, из которых были удалены семена. Споры прорастали и вызывали гибель некоторых лежащих под ними клеток в течение 24 ч. Инфекционные капельки собирали через различные интервалы времени и изучали их влияние на новые споры. После инкубации между створками боба в течение 14 ч капельки приобретали способность ингибировать прорастание спор, а через 24 ч достигался полный фунгистазис. Вещество, ответственное за фунгицидную активность, можно было экстрагировать из объединенных инфекционных капель петролейным эфиром, но химически оно не было охарактеризовано.
Впервые вещество было охарактеризовано как фитоалексин после опытов с бобами гороха и М. fructicola, в результате которых было выявлено сходство экстрактов из инфекционных капелек [46], собранных из плодов гороха и боба. Соединение, выделенное из гороха было получено в кристаллическом виде, идентифицировано как птерокарпан и названо пизатином [189]. Вскоре после этого были заново проведены опыты с бобами, в результате чего было выделено и охарактеризовано родственное птерокарпаноидное соединение, названное фазеоллином [47, 188]. История открытия этих соединений была изложена в обзоре Круикшанка [44], где указывалось, что недавно проведенная работа с другими инфицированными растениями дала сходные результаты. Поэтому сесквитерпеноидное соединение ипомеамарон из инфицированных корней батата Ipomoea batatas [107, 138], орхинол из инфицированных клубней орхидеи Orchis militaris [90] и метоксимеллеин из инфицированных корней моркови Daucus carota [39, 40], следует рассматривать как фитоалексины. Таким образом, вскоре несколько различных химических соединений было идентифицировано как фитоалексины, все они оказались относительно низкомолекулярными веществами. Предположение о том, что фунгицидные соединения каким-то очень простым способом влияют на устойчивость к болезням, было высказано еще в ранних работах с луком и возбудителем пятнистости Colletotrichum circinans. Было обнаружено, что фунгицидные соединения диффундируют в инфекционные капельки на внешней поверхности луковицы Allium cepa, устойчивого к этому грибу. Эти соединения были идентифицированы как протокатеховые кислоты [6, 144] и катехол [145]. Кроме того, было обнаружено, что они присутствуют во внешних мертвых чешуях еще до заражения. Важной отличительной чертой этой работы было исследование на наличие этих соединений У потомства, полученного при скрещивании устойчивых и восприимчивых сортов лука. Во всех исследованных вариантах была установлена зависимость между устойчивостью и наличием в чешуях упомянутых выше соединений. Это подтверждает гипотезу о том, что эти вещества являются факторами устойчивости. Аналогичные гипотезы о том, что особые ранее образовавшиеся соединения являются основой устойчивости и у других растений, редко проверялись так всесторонне, как в этом раннем исследовании.
Несмотря на большое число данных, свидетельствующих о наличии в растениях ранее образовавшихся факторов, влияющих на устойчивость некоторых видов и сортов, трудно понять, какую роль играют эти факторы в высокоспецифичных взаимодействиях между физиологическими расами паразитов и различными сортами растения-хозяина, а также в индуцированной устойчивости. Как указывалось в главе 1, инфекционный тип обычно определяется генетически обусловленным взаимодействием между расами и сортами. Хотя уже имеющиеся факторы могут играть важную роль в природе и представлять ценность как основа некоторых форм устойчивости в новых сортах, они вызывают меньший интерес, чем фитоалексины или другие вещества и процессы, возникающие после инфекции.
Фунгицидные соединения, связанные с устойчивостью, могут возникать путем образования из неактивных предшественников под влиянием ферментов, которые в здоровых тканях обычно отделены друг от друга. Так, зерновые культуры, такие, как рожь, пшеница и кукуруза Zea mays, содержат гликозиды дигидроксибензоксазолинона и глюкозидазы, которые взаимодействуют в гомогенизированной ткани с быстрым выделением фунгицидных аглюконов [99, 264, 266]. Можно было бы ожидать, что инфекция клеток зерновых культур, которая приводит к смешиванию клеточных компонентов при реакции сверхчувствительности, вызывает выделение активных соединений. Известно, что аналогичные процессы, затрагивающие различные типы соединений, могут иметь место в ряде других видов растений, как сообщалось Ингамом [111, 112]. Выделение фунгицидных соединений в результате этих процессов укладывается в рамки первоначальной концепции фитоалексинов, предложенной Мюллером и Бёргером [177, 179]. Однако, насколько это известно, соединения, которые впервые были признаны как фитоалексины, синтезируются в сверхчувствительных тканях из отдаленных предшественников, что вызывает к ним особый интерес.
Основой для предположения о том, что пизатин и фазеоллин синтезируются, а не образуются из непосредственных предшественников, служит медленное накопление их в тканях и отсутствие данных о наличии у гороха и боба каких бы то ни было простых предшественников этих соединений. Хотя все имеющиеся в растениях фунгицидные соединения могут играть определенную роль в защитной реакции организма против инфекции, наибольший интерес привлекают к себе именно фитоалексины, пути синтеза которых активируются, очевидно, после инфекции. Можно предположить наличие многих способов, с помощью которых метаболические системы хозяина и паразита могут взаимодействовать как при активации этих путей, так и при регуляции их различных энзиматических этапов. Следовательно, изучать фитоалексины следует не только потому, что они являются факторами защиты растений, но и потому, что они играют роль посредников при специфических взаимодействиях между хозяином и паразитом. Таким образом, оставшаяся часть этой главы будет посвящена фитоалексинам тех семейств, где они были обнаружены, а также некоторым общим проблемам, связанным с процессами образования фитоалексинов и локализацией в клетках мест их синтеза. Следующая глава касается их роли в устойчивости и путей, с помощью которых они могут быть вовлечены в специфические взаимодействия между растениями и паразитами.
Фитоалексины бобовых
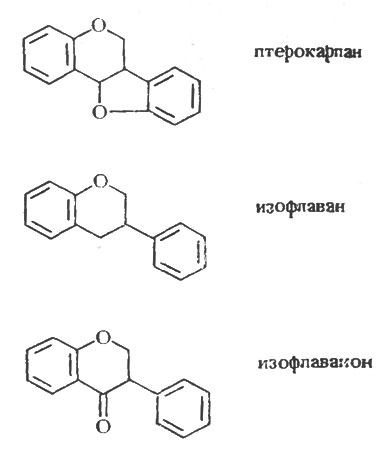
Рис. 1. Основная структура птерокарпанов, изофлаванов и изофлаванонов
Идентификация пизатина и фазеоллина как фитоалексинов в горохе и бобе стимулировало сначала попытки выделить фитоалексины из других бобовых, а затем повторное исследование ранее изученных видов на наличие в них других фитоалексинов. Оба направления исследований оказались чрезвычайно плодотворными. В люцерне Medicago sativa и сое Glycine max были обнаружены птерокарпаноидные фитоалексины; соединение, выделенное из люцерны, было названо медикарпином [233], а соединение из сои было сначала охарактеризовано как родственное фазеоллину и названо оксифазеоллин [222]. Позднее было обнаружено, что структура и название неверны, и была предложена более точная структура фитоалексина из сои [30]. В результате дальнейших исследований из бобов, помимо фазеоллина, было выделено еще три химически родственных фитоалексина: фазеоллидин [191], фазеоллинизофлаван [31] и киевитон [31, 232]. В одновременно проводимой работе было показано, что в красном клевере образуются два фитоалексина [105]; один из них идентичен выделенному из люцерны медикарпину, а другой - мааккиаин в микроколичествах образуется в инфицированном горохе наряду с пизатином [24]. В другом параллельном исследовании в люцерне бы, выявлен второй изофлаваноидный фитоалексин - сативан [13], который был также обнаружен в Lotus corniculatus наряду с родственным соединением - веститолом [22]. Медикарпин был обнаружен, кроме того, в Canavalia ensiformis [123].

Таблица 7. Фитоалексины в некоторых родах бобовых (Leguminosae)
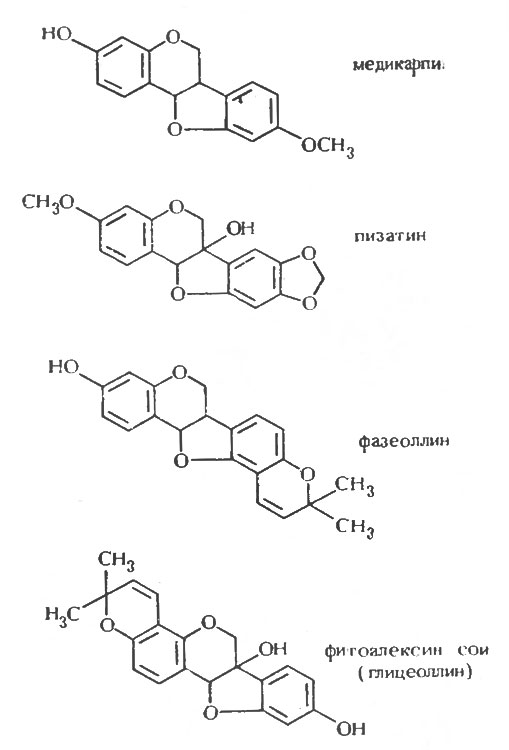
Рис. 2. Структура некоторых птерокарпаноидных фитоалексинов
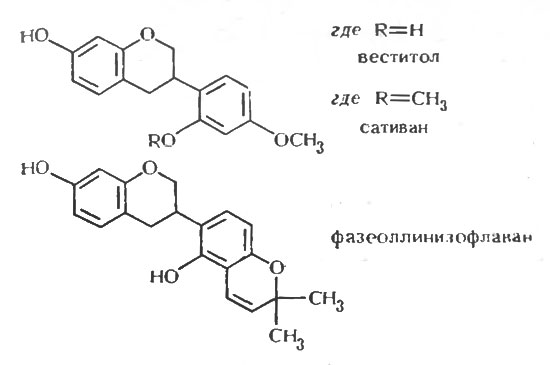
Рис. 3. Структура некоторых фитоалексинов изофлавановой природы
Таким образом, как показано в таблице 7, которая составлена в соответствии с родовой классификацией бобовых, они образуют по крайней мере один, а обычно несколько фитоалексинов [110]. Из данных, представленных в таблице, видно, что ряд бобовых образует одно и то же соединение, чаще всего медикарпин. Причина этого явления становится понятна, если рассмотреть структуру и химическое сродство этих фитоалексинов, что было сделано Ван Эттеном и Пуппке [259]. Основная структура птерокарпанов, изофлаванов и изофлаванонов представлена на рисунке 1, а структура некоторых из известных фитоалексинов, сгруппированных по соответствующим классам, - на рисунках 2, 3 и 4. Происходит ряд замещений в боковых цепях основной кольцевой структуры, и можно видеть, что медикарпин является в этом отношении одним из простейших птекарпанов. В таблице 7 также показано, что виды Vicia faba образуют два фитоалексина, названные виероном и виероновой кислотой, которые еще не были упомянуты. Виерон был впервые охарактеризован как фунгицидный компонент экстрактов из семян конского боба, которые прорастали в течение восьми дней между слоями влажной ткани. Однако Фаусетт, Фирн и Спенсер [70], хотя им и не удалось обнаружить виерон в здоровых листьях конского боба, показали, что он накапливается в них в значительных количествах после заражения листьев боба Botrytis fabae. Близкая по структуре виерону виероновая кислота была ранее обнаружена в зараженных листьях конского боба и рассматривалась как фитоалексин [143]. Сейчас ясно, что оба эти соединения должны рассматриваться как фитоалексины, потому что они образуются после заражения тканей конского боба Botrytis spp. [163] в количествах, достаточных для проявления фунгицидных свойств, и отсутствуют в здоровых тканях. По своей структуре соединения, представленные на рисунке 5, сильно отличаются от изофлавоноидных фитоалексинов других бобовых, исследованных до настоящего времени, а с фитохимической точки зрения даже противоположны им. Виерон является метиловым эфиром кетофураноидной жирной кислоты ацетиленового ряда. Эти структуры свидетельствуют о том, что у бобовых выделены фитоалексины различных типов. В настоящее время во многих видах бобовых обнаружено, как правило, несколько фитоалексинов. Возможно, что в некоторых инфицированных растениях метаболизм грибов может непосредственно обусловливать превращение одного соединения в другое, что будет более подробно обсуждаться в следующей главе. Однако в опытах Бейли и Бурдена [12] и Бейли [10] было показано, что бобы и коровий горох образуют соответственно четыре и три фитоалексина после заражения вирусом табачного некроза, который, как предполагается, сам не способен к метаболизму. Поэтому совершенно ясно, что после индукции вирусом частичного некроза в тканях растений, по крайней мере, этих видов может образоваться несколько фитоалексинов. Вопрос о том, какое значение имеет наличие часто наблюдаемой взаимосвязи между некротизацией и образованием фитоалексина, будет обсужден позднее.
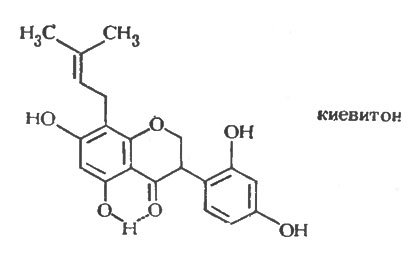
Рис. 4. Структура изофлавана киевитона, фитоалексина бобовых
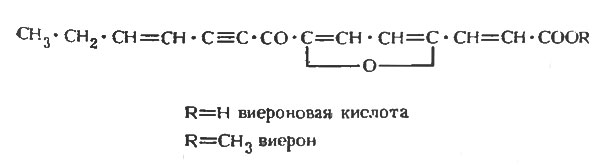
Рис. 5. Структуры кетофураноидных алифатических фитоалексинов ацетиленового ряда из конского боба Vicia faba
Фитоалексины пасленовых
Напомним, что фитоалексины, как предполагали Мюллер и Бёргер [179], образуются в клетках картофеля при развитии в них реакции сверхчувствительности к авирулентным расам Phytophthora infestans. Однако химически фитоалексин из картофеля не был выделен до работы Томиямы и др. [254], в результате которой был идентифицирован терпеноидный фитоалексин - ришитин [121], обнаруженный также в инфицированной ткани помидора [212]. Позднее из картофеля были выделены два других терпеноидных фитоалексина: любимин [176] и фитуберин [41, 263]. После инфекции в устойчивых клубнях картофеля накапливается множество других терпеноидных соединений, очевидно, в результате общей стимуляции терпеноидного синтеза в ходе реакции сверхчувствительности [139]. В перце образуется терпеноидный фитоалексин - капсидиол [84] в ответ на заражение грибом, и это же соединение образуется в листьях двух видов табака Nicotiana tabacum и N. clevelandii после заражения вирусом, вызывающим некроз листьев табака [13]. Другое родственное соединение глютинозон было выделено как фунгицидный продукт листьев другого вида табака N. glutinosa, имеющего после заражения ВТМ небольшие участки некрозной ткани [33].
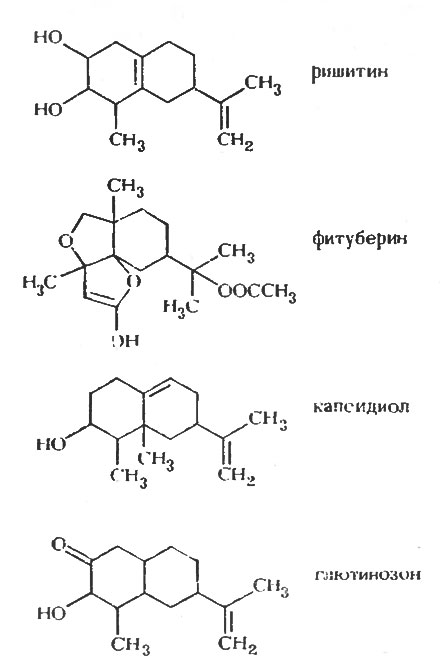
Рис. 6. Терпеноидные фитоалексины пасленовых
Таким образом, хотя у пасленовых было исследовано меньшее число видов, в этом семействе были обнаружены явления, аналогичные тем, которые наблюдались У бобовых. В отличие от изофлаваноидов родственные терпеноиды (рис. 6) встречаются у различных видов растений. Картофель и табак, исследованные наиболее интенсивно, образуют, как и бобовые, несколько активных соединений. Очевидно, и у пасленовых имеется тесная связь между образованием некроза и накоплением фитоалексина, а появление индуцированного вирусом некроза табака также приводит к образованию фитоалексина.
Фитоалексины растений других семейств
Только ограниченное число других семейств в царстве растений и несколько видов внутри них были исследованы на предмет образования в них фитоалексина; эти семейства и виды приведены в таблице 8. Замещенные изофлаваноны встречаются у одного из представителей маревых, а замещенные изокумарины - у зонтичных. Хлопчатник Gossypium barbadense из семейства мальвовых образует в ответ на инфекцию по крайней мере два нафтальдегида. Два полиеновых алифатических спирта можно считать фитоалексинами в сафлоре Carthamus tinctorius, хотя одно из этих сочинений - сафинол - можно обнаружить в небольших количествах в здоровых тканях [2]. Соединения ацетиленового ряда встречаются во многих растениях и могут обладать фунгицидными свойствами [29, 235]. Дальнейшие исследования могут выявить важность полиеновых соединений как факторов защиты у сложноцветных и других семейств. Клубни орхидей были объектом некоторых наиболее значительных ранних работ по индуцированной устойчивости, как об этом сообщалось в главе 4, в результате чего были охарактеризованы фенантреновые фитоалексины, орхинол и хирцинол в двух видах орхидных: Orchis militaris и Lorogiossum hircinum. соответственно. Решая некоторые спорные вопросы генезиса этих соединений, Гойманн и Яааг [80] и Гойманн и Керн [81, 82] показали, что в здоровых клубнях имеется незначительное количество фунгицидных веществ, но после инфекции обнаруживается высокая концентрация фитоалексинов.
| Семейство | Вид | Фитоалексин | Литературный источник |
|---|---|---|---|
| Chenopodiaceae | Beta vulgaris | 2,5-диметокси-6,7-метилендиоксифлаванон 2-гидрокси-5-метокси-6,7-метилендиоксиизофлаванон |
[83] |
| Malvaceae | Gossypium barbadense | Вергозин Гемигоссипол |
[281] |
| Umbelliferae | Daucus carota | 3-метил-6-метокси-8-гидрокси-3,4-дигидроизокумарин | [40] |
| Pastinaca sativa | Ксаитотоксин | [114] | |
| Convolvulaceae | Ipomoea batatas | Ипомеамарон | [138] |
| Compositae | Carthamus tinctorius | Сафинол | [2] |
| Дегидросафинол | [3] | ||
| Orchidaceae | Orchis militaris | Орхинол | [90] |
| Loroglossum hircinum | Хирцинол | [79] |
Из литературных данных известно только об очень небольшом числе растений, не относящихся к семейству бобовых или пасленовых, в которых образуются фитоалексины, поэтому невозможно представить себе распространенность этого процесса у высших растений. Этот недостаток информации, возможно, отражает тот факт, что другие семейства не были исследованы, а о некоторых попытках, давших отрицательные результаты, просто не сообщалось. В связи с этим следует отметить, что автору не удалось установить никакого увеличения содержания фунгицидных веществ, обнаруженных в здоровых плодах и листьях двух представителей семейства тыквенных Cucumis sativus и Cucurbita pepo, ни после заражения, ни после развития в их тканях реакции сверхчувствительности к Colletotrichum lagenarium.
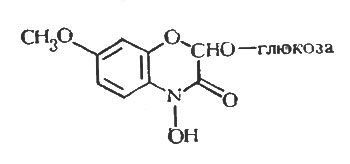
Рис. 7. Структура глюкозида дигидроксиметоксибензоксазолинона у пшеницы Triticum aestivum
Тот факт, что у тыквенных не удалось показать образования фитоалексина, используя методы обнаружения, которые легко выявляют эти соединения у бобовых, необходимо рассматривать в связи с результатами опытов автора на пшенице, представителе семейства злаковых. В этом случае использование ряда способов экстракции, а также методов разделения и биопробы, оказавшихся успешными при обнаружении фитоалексинов у бобовых, позволило выявить фунгицидное соединение в проростках пшеницы, которое обусловливало их устойчивость к стеблевой ржавчине, вызываемой Puccinia graminis f. sp. tritici. Однако свойства выделенного соединения свидетельствуют о том, что оно относится к ранее упомянутым бензоксазолинонам и, следовательно, образовалось из глюкозидного предшественника в сверхчувствительных клетках (рис. 7). Таким образом, маловероятно, что фитоалексин в пшенице может синтезироваться тем же способом, которым они образуются в растениях семейства бобовых и пасленовых. Эти два примера, приведенные при рассмотрении главного вопроса данной главы, указывают на то, что образование фитоалексина не является универсальным явлением в растительном царстве. Точно так же, как у бобовых, абсолютно разные вещества выступают в роли фитоалексинов, так и у высших растений совершенно различные механизмы защиты могут иметь место. Однако этот вопрос требует весьма интенсивного исследования, потому что сообщений о неудачах при выделении фитоалексинов, гораздо меньше, чем сообщений об удачных экспериментах.
Индукция и места образования фитоалексинов
В дискуссии о фитоалексинах у бобовых и пасленовых подчеркивалась тесная связь между появлением индуцированного вирусом некроза и образованием фитоалексинов. Возникают следующие вопросы, касающиеся механизма индуцированного синтеза фитоалексинов при различных типах инфекции и локализации этого синтеза в клетке. Является некроз причиной образования фитоалексинов или это сопутствующее явление? В каких клетках образуются фитоалексины - гибнущих, мертвых или живых? Эти вопросы не только важны для полного понимания естественной последовательности явлений в инфицированных растениях, но и имеют отношение к проблемам, связанным с повышением концентрации защитных химических веществ у растении при угрозе эпифитотий.
Вскоре после открытия пизатина, Круикшанк и Перрин [48] установили, что его образование индуцируют ионы некоторых тяжелых металлов. Затем было показано, что того же результата можно достичь с помощью широкого спектра органических веществ [9, 89, 190]. Пока не появилось удовлетворительного объяснения, посредством какого биохимического механизма индуцируется образование пизатина, но выдвинуты предположения относительно влияния их на ядерную ДНК [88]. Наиболее важным для понимания последовательности явлений, происходящих во время заражения, было обнаружение метаболита гриба Monilinia fructicola, который, присутствуя в низких концентрациях, активно способствовал образованию фазеоллина в семенных полостях створок боба [50]. Именно этот гриб, патоген мягких плодов, использовался в ранних опытах, в которых было впервые показано образование фитоалексина при развитии в клетках створок боба реакции сверхчувствительности. В результате этой работы с метаболитом гриба, названным мониликолином А, авторы пришли к выводу, что это вещество может быть ответственно за индуцирование образования фазеоллина во время инфекции. Интерес к мониликолину А еще больше возрос, когда было показано, что он может индуцировать образование фазеоллина, не убивая предварительно клетки створок боба [187]. Таким образом, можно выдвинуть гипотезу, что вещество, выделяемое паразитом, индуцирует образование фитоалексинов живыми клетками растения-хозяина. Такая гипотеза согласуется с обсуждавшейся вкратце в главе 1 концепцией, что продукт гена авирулентности активизирует защитный механизм растений, взаимодействуя с продуктом гена устойчивости хозяина. Этим защитным механизмом может быть образование фитоалексина живыми клетками. Не следует слишком уверенно приписывать молекуле, которая способна индуцировать образование фитоалексина, главную роль во взаимодействии хозяина и паразита. Так как многие органические молекулы, как было указано выше, обладают такими свойствами, не удивительно, что активные индукторы можно получить из мицелия гриба или культуральной жидкости. Необходимо выяснить, может ли индуктор действовать так же в ходе процесса зара, жения. Более веским основанием для доказательства того, что был выделен действительно индуктор, мог бы стать факт, что он обладает той же специфичностью, что и паразит, из которого он выделен. Так, специфический индуктор должен быть активным только в тканях растения-хозяина, устойчивого к авирулентной расе, образующей индуктор.
Доказательство того, что метаболиты гриба индуцируют образование фитоалексина живыми клетками, следует сопоставить с наблюдениями, свидетельствующими о том, что в ряде инфицированных растений фитоалексины начинают накапливаться примерно в то же время, когда появляются первые признаки некроза. Эти наблюдения основаны на исследованиях заражения Vicia паразитическим грибом Botrytis [162], Phaseolus паразитами Colletotrichum lindemuthianum [14, 199] и Uromyces appendiculatus [15], нескольких бобовых и Nicotiana spp. вирусами. Следовательно, образование фитоалексина, по-видимому, индуцируется гибелью клеток в результате взаимодействия хозяина и паразита.
Образование фитоалексинов удалось вызвать в двух различных экспериментах, в которых использовали физические воздействие для имитации повреждений, вызываемых обычно инфекцией. Так, постепенные физические повреждения листьев конского боба вызывали некроз и образование фитоалексина в течение 24 ч [59], в то время как сильное воздействие было неэффективно, возможно потому, что клетки погибали слишком быстро. Определенные виды повреждений, вызываемые замораживанием, также стимулировали синтез фазеоллина у боба [200], в то время как другие типы повреждений, такие, как протыкание булавкой или нанесение царапины, не вызывали образование пизатина в плодах гороха [48]. Следовательно, образование фитоалексина в процессе патогенеза могло быть обусловлено либо веществами, диффундирующими из стенок инфекционных гиф или присутствующими в них, либо процессами, связанными с гибелью клеток растения-хозяина в результате воздействия гиф.
Тот факт, что образование фитоалексина часто связано с некрозом клеток в процессе патогенеза, послужил основанием для гипотезы о том, что фитоалексины могут образовываться мертвыми или гибнущими клетками. Маловероятно, что такие клетки способны к синтезу новых молекул, хотя можно ожидать, что в них происходит гидролиз и окисление. Действительно, Ратмелл и Бендалл [902] предположили, что пероксидазы, которые, по-видимому, активны в гибнущих клетках, катализируют конечные этапы взаимопревращения изофлаваноидов, как это можно себе представить в случае образования фазеоллина. В живых клетках, расположенных вокруг мертвых, вероятнее всего, происходят более сложные синтетические процессы, причем ряд исследователей отмечали необычные свойства цитоплазмы этих клеток: повышенную чувствительность к теплу и обычно совместимым гифам [206, 224], а также повышенную плотность [175]. Обсуждая вопрос о месте синтеза фитоалексинов, уместно вспомнить о выявлении необычной флюоресценции в вакуолях живых клеток конского боба, примыкающих к некротизированным клеткам, инфицированных Botrytis cinerea [163]. Эмиссионный спектр этой флюоресценции был идентичен спектру чистых растворов виерона и виероновой кислоты. По мнению авторов, фитоалексин, по-видимому, образовывался в этих живых клетках, а не диффундировал из некротических, потому что вокруг мертвых клеток не было кольца флюоресцирующих клеток.
В настоящее время есть основания для вывода, что фитоалексины синтезируются живыми клетками. Однако нерешенным остается вопрос о факторах, стимулирующих их образование. Не вызывает сомнения, что этот процесс можно вызвать несколькими способами. Вирусные инфекции и физические повреждения могут активизировать защитную систему растения, которая, в свою очередь, индуцирует образование фитоалексина. Грибы могут выделять индуцирующие синтез фитоалексинов молекулы, высокоселективные по своему действию на клетки различных сортов растений-хозяев. Насколько тесно связан процесс отмирания клеток хозяина с образованием фитоалексинов?
Возможными кажутся несколько различных вариантов, а именно:
- (I) гибель клеток хозяина может вызывать синтез фитоалексинов в соседних клетках;
- (II) молекулы, диффундирующие из гриба, в зависимости от концентрации могут индуцировать как образование фитоалексинов, так и гибель клеток;
- (III) накопление фитоалексинов может вызывать гибель клеток хозяина, как предположили Мансфилд и др. [163].
Биосинтез фитоалексинов
Как отмечалось в начале этой главы, есть основания предполагать, что метаболические пути биосинтеза фитоалексинов в растении активизируются после инфицирования. Хотя в литературе имеется много данных относительно метаболических путей, по которым идет образование изофлаваноидных и терпеноидных соединений у некоторых организмов, активность этих путей у больных растений изучена недостаточно и совсем нет работ по биосинтезу соединений ацетиленового ряда после заражения. В результате мы не располагаем достаточным количеством данных, во-первых, чтобы подтвердить, что биосинтез фитоалексинов осуществляется из отдаленных предшественников, а не путем превращения или гидролитического расщепления из близких предшественников, и, во-вторых, чтобы оценить роль конкретных энзиматических этапов этого биосинтеза. На основе выяснения механизмов индуцированного образования фитоалексинов и оценки ключевых процессов их биосинтеза могут быть разработаны новые методы борьбы с болезнями, базирующиеся на использовании существующих в растениях систем. В этом разделе мы кратко остановимся на наиболее важных этапах биосинтеза этих соединений и приведем доказательства активизации этих этапов после заражения.
Имеющиеся в литературе данные по биосинтезу изо флаваноидов у растений были удачно суммированы Ван Эттеном и Пуппке [959]. Гризбах [85] показал, что основная скелетная структура флаваноида образована из продуктов двух метаболических циклов, а именно ацетатмалонатного и цикла шикимовой кислоты. Фенилаланин образуется через шикимовую кислоту, а затем дезаминируется ферментом фенилаланинаммонийлиазой до коричной кислоты. Коричная кислота конденсируется с ацетатом с образованием либо халькона, либо его изомера - флаванона [86]. Последующие этапы образования изофлаванов и птерокарпанов не были установлены достаточно точно, но возможные метаболические пути их образования были постулированы Ван Эттеном и Пуппке [259] на основании схемы, предложенной Вонгом [275]. Данных о наличии этих этапов у пораженных растений очень мало. Несколько исследователей показали возможность переноса меченого углерода (14С) от фенилаланина к изофлаваноидам: пизатину и фазеоллину [87, 95]. Имеется большое число данных, свидетельствующих о сильном увеличении у некоторых инфицированных растений активности фермента - фенилаланинаммонийлиазы, а Ратмелл [20] показал, что это происходит до накопления фазеоллина у боба, инфицированного Colletotrichum lindemuthianum. Не было установлено участие фермента халконфлавонизомеразы в биосинтезе фазеоллина, но заражение многих растений часто сопровождается повышением активности пероксидазы, что может играть определенную роль в образовании изофлаваноидов из непосредственных предшественников [202]. Таким образом, было установлено, что скорость биосинтеза коричной кислоты у инфицированных бобовых увеличивается, возможные последующие этапы на пути от коричной кислоты к птерокарпанам и изофлаванам изучены весьма слабо. Другой источник флаваноидов у бобовых, а именно их гликозидные предшественники, был предложен Олахом и Шервудом [186], которые обнаружили, что в листьях люцерны после заражения Ascochyta значительно увеличивается активность гликозидаз, возможно, грибного происхождения. Однако Олах и Шервуд [185] показали, что концентрация флаваноидных глюкозидов в больных листьях при этом не уменьшалась, как можно было ожидать, если бы имел место гидролиз. Необходимо дальнейшее исследование метаболизма пораженных листьев люцерны, чтобы найти источник появления в них фитоалексинов, медикарпина и сативана, и выяснить, происходит ли активация синтеза флаваноидных гликозидов после инфекции, что, по-видимому, необходимо постулировать для объяснения наблюдаемых явлений. На основании результатов аналогичных опытов Ратмелл [201] был вынужден сделать вывод, что заражение боба специфически стимулировало синтез изофлаваноидов, так как ему не удалось выявить никаких одновременных изменений во флаваноидах, образовавшихся из гликозидов.
Считается, что терпеноиды синтезируются в растениях из ацетата и мевалоната, и были получены доказательства преимущественного переноса изотопа 14С из этих предшественников в ришитин и родственные соединения при развитии реакции сверхчувствительности в дисках из клубней картофеля [218]. Ших, Куч и Уильямс [219] высказали интересную мысль о том, что переключение послеинфекционного метаболизма инфицированной ткани картофеля зависит от совместимости инфицирующего организма и растения-хозяина и направлено на накопление терпеноида в устойчивых тканях или на образование стерогликоалкалоида в восприимчивых. Эта идея будет обсуждаться в следующей главе. Огуни и Уритани [184] охарактеризовали путь биосинтеза ипомеамарона из ацетата и мевалоната в инфицированным клубнях батата и частично исследовали роль дегидроипомеамарона как предшественника фитоалексина. Таким образом, имеются достаточно убедительные данные по усилению биосинтеза терпеноидов в инфицированных растениях, хотя пока еще невозможно идентифицировать ключевой этап этой активации.
Заключение
Получено огромное количество материалов, подтверждающих существование у растений некоторых сeмейств фитоалексинов, характеризующихся различной химической структурой. В следующей главе будет обсуждаться доказательство участия фитоалексинов в регуляции развития гриба при определенных типах взаимоотношения хозяина и паразита. Необходимо еще ответить на некоторые важные вопросы, касающиеся способов, с помощью которых индуцируется образование фитоалексинов после инфекции, и получить данные, подтверждающие биосинтез фитоалексинов из отдаленных предшественников. Понимание этих процессов может помочь найти новые возможности защиты растений от болезней путем управления естественно происходящими процессами. Однако нельзя делать вывод об универсальности фитоалексинов в растительном царстве, так как высшие растения, возможно, используют несколько различных способов химической защиты.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'