Глава 10. Солнечные комбинаты и химические перпетуум-мобиле (Функция растительных сообществ)
Понять функцию фитоценоза в отрыве от прочих компонентов экосистемы практически невозможно, так как большинство связей в растительном сообществе проходит либо через среду, либо определяется гетеротрофными компонентами - животными и микроорганизмами.
Тем не менее будет полезным сказать хотя бы несколько слов о функции растительного сообщества и ее месте в функции экосистемы, памятуя, что из двух ее основных сторон - фиксации солнечной энергии и кругооборота минеральных элементов, воды, углекислого газа и кислорода - первая еще в какой-то мере автономна, а вторая целиком зависит от деятельности гетеротрофных компонентов экосистемы.
Сообщество растений - это истинный солнечный комбинат: на каждый сантиметр поверхности Земли ежегодно падает гигантское количество энергии - в среднем около 55 ккал солнечной энергии, но лишь сравнительно небольшая часть этой энергии (в зависимости от благоприятности остальных условий она может варьировать в интервале от 1 до 0,05%) связывается растениями в результате фотосинтеза в органическое вещество, из которого состоит тело растения. Существование сложной структуры с дифференцированными в отношении светового ресурса нишами ценопопуляций обеспечило солнечным комбинатам из сосудистых растений преимущество по сравнению с растительностью мезозоя, которую формировали бессосудистые голосеменные, плауны и хвощи.
В лесах с развитыми ярусами подлеска, кустарников и трав общая площадь листовой поверхности на каждый 1 м2 почвы составляет 4-6 м2, причем в этой массе содержится до 4 г хлорофилла. В среднем 1 г хлорофилла связывает в год около 1000 ккал солнечной энергии, что соответствует примерно 0,5 кг сухого органического вещества.
Леса занимают около 10% площади земной поверхности, но в силу насыщенности хлорофиллом фиксируют примерно половину солнечной энергии, если учесть всю фиксацию биосферы, включая и океан. Другие типы сообществ менее продуктивны, так как имеют меньшую площадь листовой поверхности и меньшее содержание хлорофилла.
То органическое вещество, которое является результатом фотосинтеза, если исключить из него неизбежные расходы на дыхание (а эти расходы составляют не менее половины от общей связываемой растениями солнечной энергии), называется чистой первичной биологической продукцией. Чистую биологическую продукцию, которая измеряется как количество сухого органического вещества (или в пересчете на углерод, или при выражении в единицах энергии-калориях), накопленного на единицу площади за единицу времени (обычно ее измеряют в граммах на 1 м2 за 1 год), нельзя путать с биомассой. Биомасса - это то количество органического вещества, которое может быть учтено в надземной и подземной частях сообщества в момент наблюдения и представляет собой для многолетников сумму чистой биологической продукции за весь период жизни за вычетом потерь, связанных с питанием животных и отмиранием части листьев, стеблей и корней. Биомасса равна чистой биологической продукции только у однолетников, отношение биомассы и чистой биологической продукции у многолетников меняется в более широких пределах, чем продуктивность. В лесах этот показатель может достигать 50, а в луговых и степных сообществах - 1-3.
Чистая продукция меняется от 3000 г/м2/год в тропических лесах практически до нуля в экстрааридных пустынях, лишенных растений, или в условиях Антарктиды с вечными льдами на поверхности суши, а запас биомассы соответственно - от 60 кг/м2 до 0. Уиттекер предлагает по продуктивности делить все сообщества на четыре класса.
- Сообщества высшей продуктивности, 3000-2000 г/м2/год. К этой группе относятся тропические леса, посевы рпса и сахарного тростника. Запас биомассы в этом классе продуктивности, как и во всех прочих случаях, разумеется, очень различен и превышает 50 кг/м2 в лесных сообществах и равен продуктивности у однолетних сельскохозяйственных культур.
- Сообщества высокой продуктивности, 2000-1000 г/м2/год. В эту группу включены листопадные леса умеренной полосы, луга при удобрении и посевы кукурузы. Максимальная биомасса приближается к биомассе первого класса, минимальная соответственно равна чистой биологической продукции однолетних культур.
- Сообщества умеренной продуктивности, 1000-250 г/м2/год. К этому классу относится основная масса возделываемых сельскохозяйственных культур, степи, кустарники. Биомасса степей меняется в пределах 0,2-5 кг/м2.
- Сообщества низкой продуктивности, ниже 250 г/м2/год,- пустыни, полупустыни (в отечественной литературе их называют чаще опустыненными степями), тундры.
Нетрудно заметить, что солнечный свет ни в коей мере не лимитирует продуктивность растительных сообществ, так как всегда находится в избытке. Продуктивность в первую очередь зависит от количества воды, так как на производство 1 г сухого органического вещества растения в большинстве своем затрачивают 700-1000 г воды. Исключение составляют кактусы, приспособившиеся расходовать воду более экономно - за счет уже упомянутого кислого метаболизма фотосинтеза. Однако одна вода при отсутствии элементов минерального питания и тепла не может быть фактором повышения урожайности. Именно полное обеспечение потребностей растений водой и элементами минерального питания при достаточном количестве тепла позволяет достигать каждому виду и соответственно сообществам предельно высоких значений продуктивности. В этом отношении показателен опыт организации орошения на посевах трав в нашей стране: наилучшие результаты - до 150 ц сухой массы на 1 га (что соответствует 1500 г/м2/год чистой биологической продуктивности только в надземной массе, а при учете подземной массы это число, видимо, должно быть удвоено) - дает полив трав при многократном дробном внесении удобрений в условиях южных районов страны - Волгоградской обл., Казахстана и т, д. По мере убывания тепла эффективность полива падает и окупаемость затрат на полив по ряду степная зона-лесостепь-лесная зона соответствует ряду 3-2-1.
Первичная биологическая продуктивность является фундаментом жизни на Земле, основанием энергетической пирамиды. Примерно 10% этой продукции связывают животные-консументы из числа травоядных млекопитающих, птиц, поедающих плоды, насекомых и т. д. Остальные 90% разлагаются сапротрофами- микроорганизмами и грибами или поступают на более или менее длительное хранение в запасы мертвого вещества, называемого детритом. Кратковременно существующими формами детрита являются опадающие мертвые листья, длительно существующими - гумус, торф, сапропель (органические отложения дна озер). Травоядные консументы служат нищей плотоядных первого порядка, которых последовательно поедают плотоядные второго, третьего и даже четвертого порядка. Каждый раз, когда в этой последовательности трансформации энергии, фиксированной растительным сообществом (пищевой цепи), происходит повышение уровня, рассеивается примерно 90% энергии. Таким образом, если допустить, что сообщество фиксировало 1% солнечной энергии, то из 10% потребленного органического вещества растениями (т. е. 0,1% усвоенного солнечного света) травоядные фиксируют также 10% и остаток солнечной энергии в его теле уже соответствуют только 0,01%. Хищник, питающийся травоядными, получает лишь 0,001%, более крупный хищник второго порядка - 0,0001, еще более крупный хищник (например, тигр) - 0,00001%. На рис. 16 показана схема трансформации энергии в экосистеме.
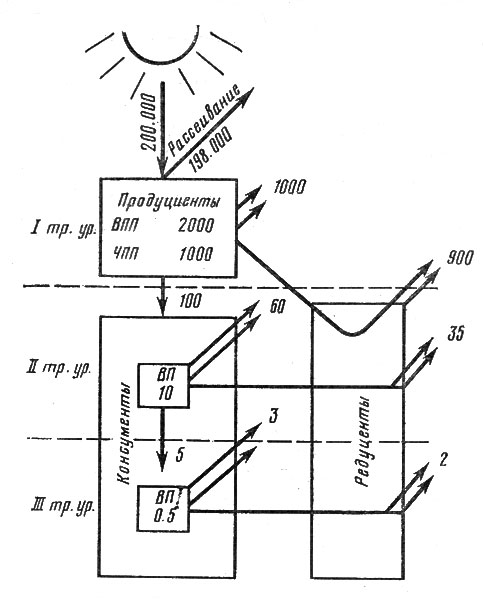
Рис. 16. Схема, иллюстрирующая энергетику сообщества и экосистемы. Размерность ккал/м2/полгода; ЧПП - чистая первичная продукция; ВПП - валовая первичная продукция; ВП - вторичная продукция
Понятно, что система функционирует в течение шести месяцев и из количества падающей за это время энергии ею усваивается 1% при фиксации в ЧПП 0,5%. Плотоядные поедают 10% чистой продукции, а остальное разрушают редуценты, причем вся энергия в этом случае теряется при их дыхании. Из энергии, полученной травоядными, 10% тратится на рост и размножение, а остальное - на дыхание и экскрецию, продукты которой служат пищей редуцентам, которые также сжигают их в процессе дыхания. В свою очередь, хищники, поедающие плотоядных, часть полученной энергии затрачивают на дыхание, а остальное (экскременты и их тела после смерти) становится пищей редуцентов, возвращающих остаток энергии во внешнюю среду. Таким образом, в этой простейшей схеме, где отсутствует блок "детрит" и число трофических уровней ограничено тремя, можно уяснить принцип стабильной экосистемы: приход энергии и ее расход в процессе дыхания организмов всех блоков одинаковы. Двойными стрелками показаны затраты на дыхание.
Закономерности трансформации энергии наводят на серьезные размышления об использовании продуктивности биосферы человеком. Людям свойственно смешанное питание - они потребляют продукцию и первого уровня (хлеб, овощи, фрукты и т. д.) и второго (мясо, молоко). Растительная пища несравненно дешевле, чем продукция животных, которую называют вторичной биологической продукцией. Расчеты показывают, что количество энергии, необходимое для откорма одной коровы, эквивалентно 1,5 т бензина. Поскольку продукция животных важна для человека потому, что является основным источником белка, во всем мире происходит интенсивный поиск возможности замены животного белка на более дешевый белок высокобелковых бобовых культур, таких, как соя, бобы, фасоль, арахис и т. д.
Продуктивность океана по сравнению с продуктивностью суши несравненно ниже и в среднем соответствует продуктивности пустынь. Исключение составляют лишь прибрежные зоны, т. е. береговые шельфы, где солнечный свет достигает поверхности дна и потому продуктивность может подниматься до уровня третьего класса и реже (области коралловых рифов и эстуариев) второго и даже первого класса. Общий запас биомассы растений океана примерно в тысячу раз ниже, чем запас биомассы растений суши. 90% биомассы суши сконцентрировано в лесах.
Солнечный свет дает сообществу растений энергию для продуцирования органических веществ, т. е. для связывания элементов минерального питания, углекислого газа и кислорода в соединения, потребляя которые, могут жить животные (в том числе и человек) и сапротрофы. В последние годы на эту способность растений все чаще обращают внимание энергетики. Уже появились проекты электростанций на солнечной энергии, которая не непосредственно концентрируется системой зеркал, а накапливается зелеными растениями. В качестве накопителей предполагается использовать не традиционную древесину, а одноклеточные водоросли, которые быстро растут, делятся и отмирают. Сжигая массу высушенных мертвых водорослей и возвращая золу в водоем - накопитель энергии, можно создать замкнутое водное сообщество, продукция которого преобразуется в наиболее приемлемую для промышленности энергетики форму электроэнергии.
Есть проекты использования в качестве горючего растительного масла и спирта, получаемых тут же, в хозяйстве, которое, таким образом, обретает энергетическую автономность.
Подобно тому как хлорофилловое зерно является входом потока энергии, усваиваемого растительным сообществом и далее протекающего через экосистему по пищевым цепям, корни растений открывают поток питательных элементов, который, как в перпетуум-мобиле, циркулирует из почвы в сообщество и возвращается обратно. Естественно, что этот вариант "вечного двигателя" далек от того, который пытались создать средневековые схоласты. Источник движения "химического перпетуум-мобиле" лежит вне системы - это энергия солнца. Корни растений разветвлены на тончайшие окончания, но поверхность всасывания во много раз увеличена за счет гифов грибов, которые обволакивают корни и формируют микоризу, один из эволюционных феноменов симбиоза, обеспечивающий растениям возможность интенсивного поглощения элементов минерального питания. Далее питательные вещества текут в стебли и листья. У деревьев максимум питательных веществ в листьях и минимум - в древесине. Поскольку листья ежегодно (или раз в несколько лет у вечнозеленых растений) опадают, то вновь синтезированные органические вещества с помощью сапробов (микроорганизмов, грибов и др.) разрушаются до простейших форм и включаются в новый цикл поглощения. Кроме возврата элементов минерального питания в почву с опадом, существенную роль играет и выщелачивание (смыв) листьев дождевыми водами, причем для натрия и калия возврат в почву через смыв больше, чем с опадающими листьями. Возвращаются в почву и вещества, фиксированные в древесине, которая подвергается действию гнилостных сапрофитов, однако этот цикл оказывается несравненно более медленным.
Количество питательных веществ, возвращаемых в почву, зависит в первую очередь от чистой первичной продукции сообщества, а скорость его повторного включения в циклы обмена - от типа сообществ и климата. В теплом тропическом лесу мягкие листья разрушаются сапрофитами в течение нескольких месяцев, но жесткие иголки ели сохраняются в холодной тайге в неразрушенном состоянии годами. Разные элементы минерального питания обращаются в сообществе с разной скоростью. Для листопадных лесов умеренной полосы характерна следующая последовательность по быстроте обращаемости элементов: сера, калий, марганец, азот, железо, кальций, фосфор, углерод.
Кроме скорости оборота в сообществе, каждый элемент питания имеет еще и структуру запаса, т. е. соотношение фракций, распределенных в почве, подстилке и непосредственно в растениях сообщества. В лесу нитраты и фосфаты, к примеру, сконцентрированы в основном в растениях, а кальций и натрий - в почве. Чем большая доля элемента фиксирована в растении по сравнению с почвой, откуда он может быть поглощен гифами грибов или непосредственно поверхностями корней, тем компактнее кругооборот элемента в сообществе.
Сообщества разных экологических условий отличаются количествами элементов, вовлекаемых в кругооборот, и общей компактностью кругооборота. Именно сообщества являются основными "инструментами", при помощи которых климат перерабатывает материнскую породу в почву. Если сравнить такие контрастные сообщества, как тропические леса, тайга и степь, то наличие в первых двух случаях большого количества воды обусловливает выщелачивание элементов в глубь почвы. Однако быстрый распад мягких листьев тропических деревьев продуцирует в основном углекислоту, а медленный распад хвои - полифенольные соединения.
Тропические почвы способны аккумулировать железо, привносимое нисходящими токами воды. Этот почвенный материал под воздействием воздуха затвердевает и образует слой, подобный скальным породам,- буквально сцементированный горизонт вмыва. Напротив, под хвойными породами в таежных лесах из верхних горизонтов почвы выщелачивается большинство минеральных соединений. Относительно малоподвижен лишь кремний, благодаря которому и возникает белесый цвет одного из верхних горизонтов почвы. Два разных типа сообществ продуцируют, таким образом, тропическую латозольную в одном случае и таежную подзолистую почву - в другом.
Степи распространены в условиях, где избытка осадков нет, и потому там практически не происходит выщелачивания. Травы степи содержат питательных веществ намного больше, чем лесные растения. Если свежий детрит леса - подстилка - составляет не более 1 % от биомассы сообщества, то аналогичный детрит степи, называемый ветошью, может превышать (особенно, если режим степи заповедный и отсутствуют пасущиеся животные) 50% биомассы. В этих условиях формируется чернозем - наиболее богатая почва с высоким содержанием питательных веществ. Общая компактность кругооборота, таким образом, падает по ряду тропический лес-тайга-степь.
Кругообороты элементов питания разных сообществ тесно связаны друг с другом через посредство надземного и внутрипочвенного стока и через атмосферу. В особенности тесно взаимопереплетены циклы элементов сообществ водосборов, т. е. территорий, имеющих общий сток в реку или озеро. Если в районе водосбора растительность лесного типа не нарушена, то значительная часть элементов фиксируется в сообществах и почвах, вынос их ограничен и обычно компенсируется за счет притока элементов, поставляемых процессом выветривания и также атмосферными осадками (дождевая вода далеко не так чиста, как принято думать).
Если же водосбор нарушен, то вынос элементов возрастает. Описанные Уиттекером в 1980 г. исследования водосбора Хаббард Брук, часть которого была покрыта лесом, а часть подверглась рубке, показали, что отсутствие сообществ, способных консервировать запас питательных веществ, резко увеличило их вынос в реку. Так, количество выносимого кальция увеличилось в 8 раз, калия - в 15, а нитратного азота (за счет активизации деятельности окисляющих микроорганизмов) - в 30 раз.
Через водные артерии кругообороты водосборов связаны с кругооборотами веществ в океанических экосистемах, наиболее продуктивные части которых - эстуарии и литорали - обязаны своим существованием именно приносу элементов минерального питания. Океан, в свою очередь, частично возвращает эти элементы с осадками, которые содержат соли, поднятые вверх брызгами воли и растворившиеся впоследствии в капельках воды облаков.
Таким образом, циклы питательных элементов внутри отдельных биогеоценозов, соответствующих сообществам, так же как и циклы энергии, в конечном итоге сплетаются в единый биогеохимический цикл экосистемы высшего порядка - биосферы.
Купить гидрозамок на сайте https://www.gidro.ru/catalog/gidroklapannaya_apparatura/gidroz../.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'