Создание новых растений
Селекция растений
Высокие нынешние урожаи сельскохозяйственных культур стали возможны в значительной мере благодаря искусству селекционеров, которым удалось создать немало продуктивных болезнеустойчивых сортов. Один из выдающихся успехов был достигнут с кукурузой. Современная кукуруза - это гибрид, соединивший в себе гены от нескольких инбредных линий. Генетикам было давно известно, что длительный инбридинг понижает мощность и продуктивность большинства сельскохозяйственных культур и домашних животных. Скрещивание же некоторых высокоинбредных линий нередко вызывает явление, называемое гетерозисом, или гибридной силой. Потомки от таких скрещиваний гораздо крупнее и отличаются от родительских сортов большей мощностью и продуктивностью, очевидно, вследствие наличия в их клетках особых сочетаний генов. Несмотря на новейшие сведения, полученные в области молекулярной биологии, мы не можем дать точного биохимического объяснения этому усиленному росту гибридов.
Для создания современной гибридной кукурузы были использованы четыре отдельные линии. Сначала эти линии скрещивали попарно, а затем два полученных таким образом гибрида скрестили между собой и в результате этого "двойного скрещивания" получили гибридную кукурузу (рис. 16.1). Разумеется, семена, производимые такими растениями при самоопылении, не "размножаются в чистоте", а отличаются высокой изменчивостью. Поэтому фермерам приходится ежегодно покупать новые семена, производимые на специальных семенных фермах. Несмотря на такие дополнительные расходы, американские фермеры идут на это, поскольку при достаточном внесении удобрений затраты на семена окупаются повышением урожайности. Прибыли, полученные в результате повышения урожайности за счет выращивания одной лишь гибридной кукурузы, могли бы покрыть всю сумму, затраченную на изучение растений в США за все время существования страны. Первоначальное повышение урожаев кукурузы в 30-х годах и позже было обусловлено введением этих генетических гибридов, а дальнейшее их повышение связано с возрастающим применением химических удобрений, а затем и пестицидов. Гибридные сорта выведены теперь и для многих других растительных культур,, и, несмотря на высокую стоимость производства гибридных семян, именно эти семена обеспечивают почти по всему земному шару богатые урожаи и хорошие доходы.
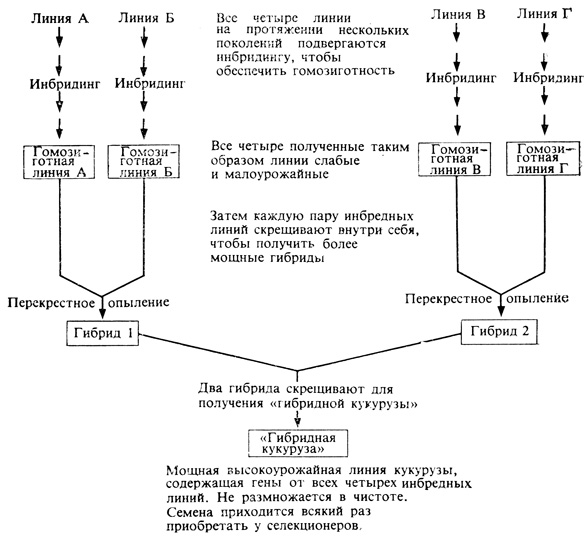
Рис. 16.1. Современная 'гибридная кукуруза' была создана путем постепенного объединения генетического материала четырех различных инбредных линий. Свойства конечного сорта, естественно, зависят от генетической природы исходных инбредных линий
При помощи отбора и скрещивания также можно повысить урожайность культур, создавая сорта, приспособленные к особым местным условиям. Многие старые местные сорта, издавна выращиваемые в данной области, хотя и малоурожайны, при внесении в почву небольших количеств удобрений оказываются более продуктивными, чем высокоурожайные сорта, созданные для интенсивного земледелия; к тому же они более устойчивы к заболеваниям. Подобные сорта содержат ценный генофонд, ожидающий того, чтобы его начали использовать для областей с экстремальными условиями.
В настоящее время селекционеры уделяют особенно много внимания поискам солеустойчивых растений для выращивания их в областях, почвы которых стали засоленными (см. также гл. 14). Некоторые дикорастущие виды томатов, например, произрастают в приморских районах, куда доходят соленые брызги океана. Гены, имеющиеся у этих видов, могли бы повысить солеустойчивость культурных сортов томатов. Проводятся также испытания на солеустойчивость хранящихся в семенных банках хлебных злаков; при этом уже удалось выявить у них значительные различия, которые могут послужить основой для выведения солеустойчивых сортов. Это показывает, какую важную роль играют семенные банки, даже если в них хранятся "устаревшие" сорта. Аналогичные программы проводят в поисках холодо- или засухоустойчивых сортов.
Изменение старых сортов растений с помощью новых методов
Скрещивания между существующими растениями, конечно, не исчерпывают всех потенциальных возможностей получения новых сочетаний геномов. Обычные скрещивания следует продолжать не только для повышения урожайности сельскохозяйственных растений и их качества, но также для их защиты от новых рас микроорганизмов и насекомых-вредителей. Кроме того, необходимо развивать дальше новые методы, в основе которых лежат использование тканевых и клеточных культур и соматическая гибридизация путем слияния протопластов, хотя все это остается пока еще лишь на уровне любопытных лабораторных экспериментов. Недавно в результате слияния протопластов был получен настоящий соматический гибрид между картофелем и томатом.. При выращивании в культуре таких гибридных клеток в начале образовывалась масса недифференцированной ткани (каллус), а затем и целые растения с некоторыми признаками обоих родителей. Чтобы доказать гибридную природу этих растений, из них выделяли и анализировали рибулозобисфосфат-карбоксилазуфермент цикла Кальвина, состоящий из больших субъединиц белка, определяемых генами, содержащимися в хлоропластах, и малых субъединиц, определяемых ядерными генами. Было показано, что у гибрида большие и малые белковые субъединицы происходят от разных родительских особей. Это первый случай, когда путем соматической гибридизации было создано растение, которое нельзя (или во всяком случае не удалось до сих пор) получить с помощью обычных методов полового скрещивания. Метод соматической гибридизации в ближайшем будущем преподнесет нам, несомненно, немало других неожиданностей.
Для экспериментов по трансформации растительной клетки наиболее подходящим объектом, почти наверное, окажутся протопласты. Мы уже располагаем рядом факторов, способных переносить гены из одной клетки в другую; самым многообещающим, из таких переносчиков служит Ti-(tumor inducing) плазмида Agrobacterium tumefaciens (возбудителя рака корневой шейки). Эту небольшую кольцевую молекулу ДНК, обусловливающую превращение нормальных клеток в клетки раковой опухоли, можно "надрезать" и разомкнуть, используя рестрикционные эндонуклеазы, выделяемые из различных микроорганизмов (рис. 16.2). К разомкнутому таким образом кольцу плазмидной ДНК можно добавить другие фрагменты ДНК, полученные аналогичным образом из клеток, гены которых подлежат переносу. В присутствии соответствующих источников энергии и ферментов, называемых лигазами, можно вызвать соединение разомкнутых концов молекул ДНК. В некоторых случаях при этом получаются гибридные плазмиды, в замкнувшееся кольцо которых включаются желаемые гены эукариотических клеток. Такие плазмиды можно затем использовать для заражения протопластов растения-хозяина, в которое предполагается перенести эти гены. В случае удачи эксперимента плазмида проникает в клетку и реплицируется вместе с хромосомной ДНК. Части плазмидной ДНК могут затем высвободиться из кольцевидной молекулы и включиться в геном-рецептор, превратившись в постоянную часть генетического аппарата теперь уже измененного растения-хозяина. Предлагалось много планов для проведения экспериментов по переносу генов, определяющих фиксацию азота, устойчивость к возбудителям болезней, более высокую аминокислотную продуктивность и более мощный габитус. Хотя в настоящее время эти планы кажутся несколько фантастичными, их следует иметь в виду на будущее. Генная инженерия почти несомненно будет играть большую роль в сельском хозяйстве XXI в.
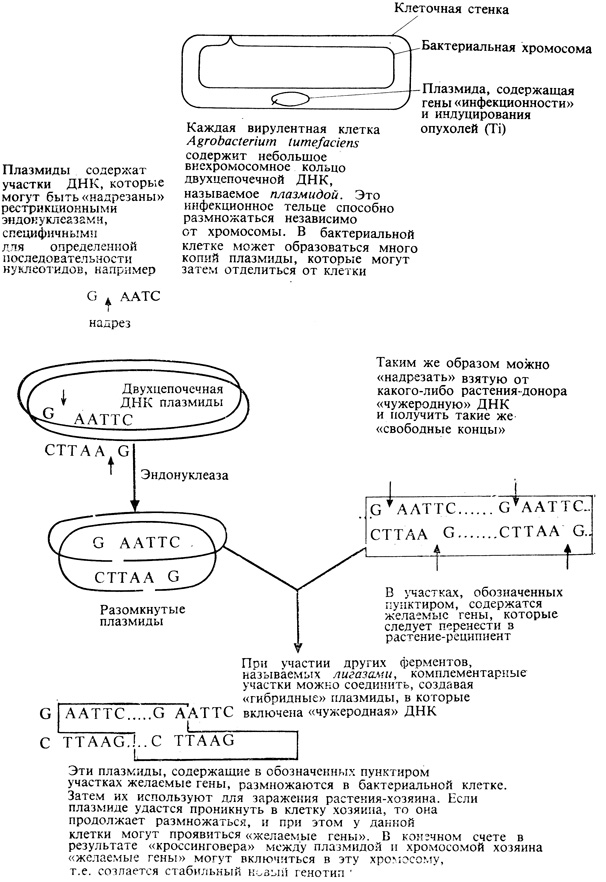
Рис. 16.2. Используя эндонуклеазы и бактериальные плазмиды, можно клонировать гены высших растений, а быть может, и вводить их новым растениям-хозяевам
Протопласты, полученные из листьев картофеля, уже используются для отбора вариантов, наиболее устойчивых к неблагоприятным воздействиям и к болезням (рис. 16.3). Создается впечатление, что каждый лист представляет собой мозаику из клеток, различающихся по своему генетическому составу; эти различия могут быть обусловлены генными мутациями, такими хромосомными изменениями, как делеции, или неравномерным распределением плазмид при клеточном делении. В чем бы ни состояла причина генетического изменения, эти клетки можно выделять в виде протопластов, из которых в результате регенерации сначала вновь образуются клетки, а затем каллусы и целые растения; полученные таким образом экземпляры можно исследовать, с тем чтобы выявить имеющиеся между ними различия. Этот мощный метод, очевидно, найдет широкое применение в будущем.
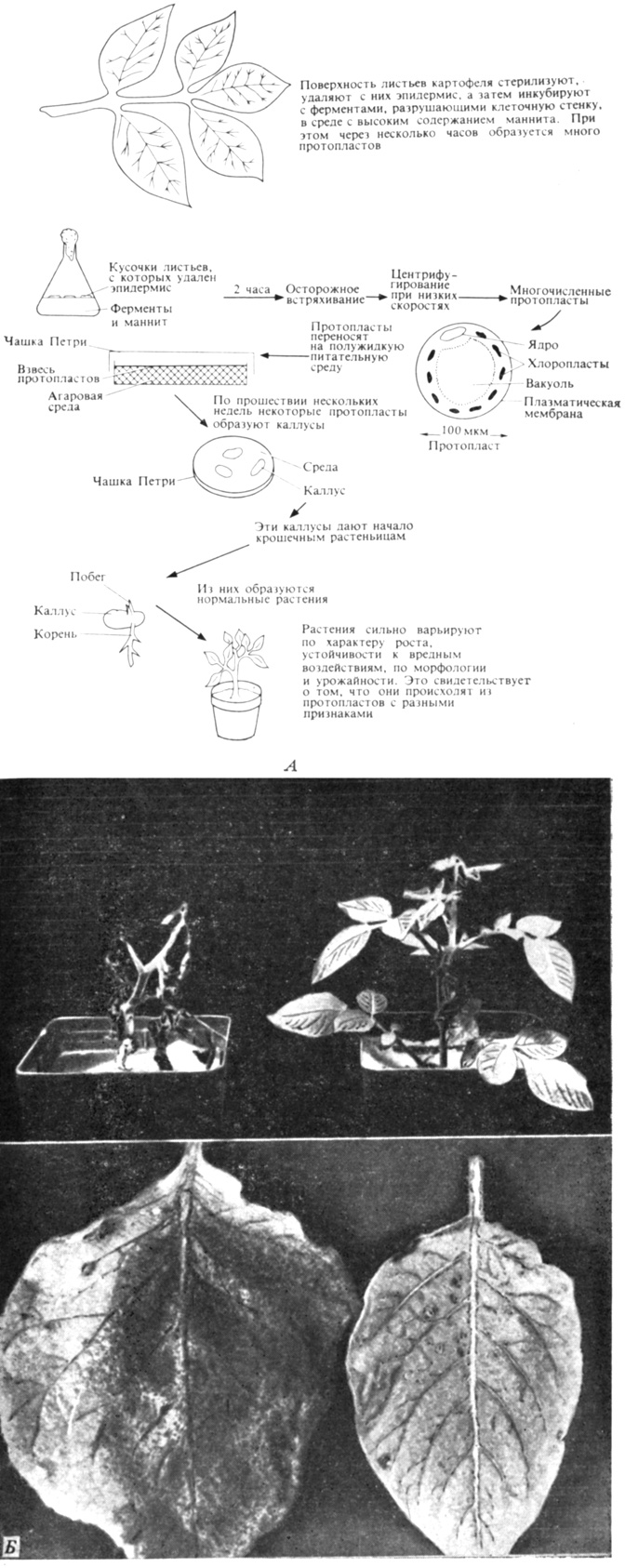
Рис. 16.3. Протопласты, выделенные из листьев картофеля, можно использовать для регенерации целых растений (А). У таких растений наблюдается высокая изменчивость (Б), что указывает на изменчивость клеток (протопластов), из которых они возникли
Близкий метод - культура клеток в присутствии антиметаболитов (рис. 16.4). Следует ожидать, что любая клетка, выращиваемая в присутствии антиметаболита, будет производить избыточное количество нормального метаболита. Если такую клетку выращивать в культуре, то она, возможно, даст начало растению, продуцирующему желаемый метаболит в больших количествах. Такой эксперимент был успешно проведен с клетками моркови. При введении в среду 5-метилтриптофана (5-МТ) большинство клеток гибнет, поскольку 5-МТ - эффективный антагонист триптофана. Но некоторые клетки при этом не гибнут; они, как оказалось, содержат большие количества эндогенного триптофана, потому что имеющийся у них фермент - антранилатсинтетаза, участвующая в образовании индольного кольца триптофана, - не подавляется, как это обычно бывает, конечным продуктом реакции. В результате этот фермент обусловливает "избыточную продукцию" триптофана, а растения, регенерированные из таких отобранных клеток, богаче триптофаном, чем те линии, из которых они происходят. Этот метод, несомненно, будет использоваться в будущем для создания линий важных сельскохозяйственных растений с повышенным содержанием витаминов, аминокислот, стероидов и других желательных продуктов, вырабатываемых растениями.
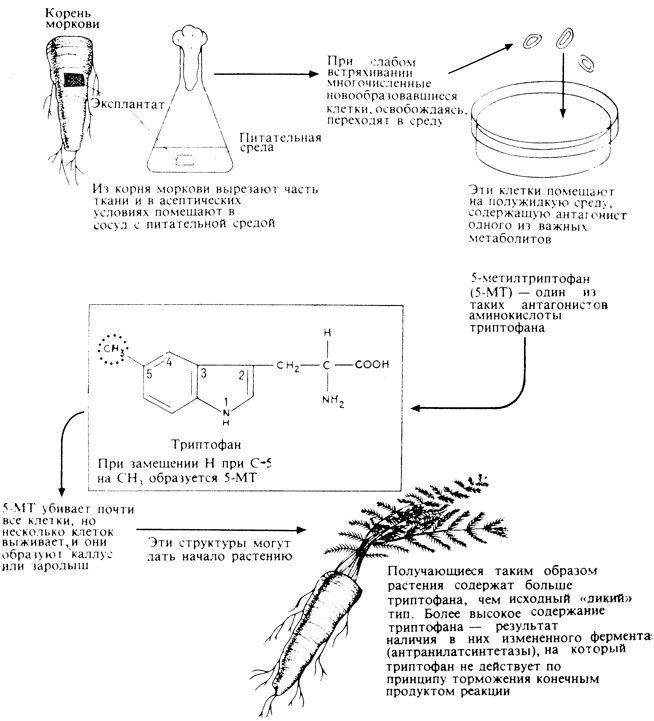
Рис. 16.4. Нормальную изменчивость клеток можно использовать для отбора
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'