Как действует фитохром?
Поскольку фитохрома в растительных клетках очень немного и малые количества энергии, поглощенные этим пигментом, могут вызывать большие физиологические сдвиги, очевидно, что первичное поглощение фотонов должно быть связано с клеточной реакцией каким-то механизмом усиления. Такой механизм мог бы включать управление функцией генов, активностью ферментов, свойствами мембран или содержанием таких веществ, как гормоны, которые сами эффективны в ничтожных количествах. Все эти процессы, видимо, имеют место в определенных ситуациях, но, судя по новейшим данным, "основным" процессом, который может приводить ко всем остальным, возможно, является регуляция функций мембран.
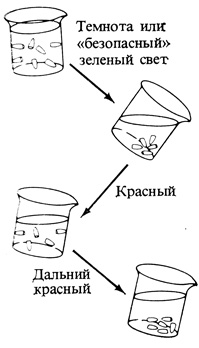
Рис. 11.17. Отрезанные кончики корней ячменя, выращенного в темноте, погружают в специальный раствор в стеклянном стакане, внутренняя поверхность которого обработана так, что она несет отрицательный заряд. Когда стакан вращают, а затем наклоняют, кончики корней при воздействии только 'безопасным' зеленым светом опускаются на дно, а при воздействии красным светом прилипают к боковой поверхности. Дальний красный свет изменяет эффект красного света на противоположный. (По данным Tanada. 1968. Proc. Natl. Acad. Sci., 59, 376-380.)
Онтогенетические реакции, такие как инициация цветения, прорастание семян и деэтиоляция, несомненно, связаны с радикальными сдвигами в химизме, структуре и функции растительных клеток. Эти сдвиги в свою очередь зависят от изменения активности многих ферментов, а также от синтеза новых ферментов. Так как ферменты представляют собой белки и их синтез определяется процессами трансляции и транскрипции, состояние фитохрома должно влиять или на какой-то один из этих процессов, или на оба. Мы не знаем, как фитохром осуществляет это влияние. Он мог бы связываться с ядерным хроматином, оказывая таким образом прямое воздействие на синтез РНК и белка; его влияние могло бы быть и более тонким, возможно, связанным с изменениями в компартментации ионов внутри клетки и как следствие - в синтезе белка. Однако контроль белкового синтеза - не единственный способ действия фитохрома, так как многие регулируемые фитохромом процессы не зависят от синтеза белка и осуществляются слишком быстро.

Рис. 11.18. Листья Mimosa pudica через 30 мин после перенесения в темноту вслед за обработкой белым светом высокой интенсивности с последующим 2-минутным воздействием красного и дальнего красного света. Листья остаются раскрытыми, если последним было облучение дальним красным светом (вверху)у и складываются, если последним был красный свет (внизу). (Fondeville et al. 1906. Planta, 69, 357-364.)
Как известно, растительные гормоны быстро перемещаются по растению и эффективны в очень малых количествах. Последствия превращений фитохрома можно было бы связать с синтезом или разрушением гормонов или же с их высвобождением или связыванием. По-видимому, любой из этих процессов мог бы происходить достаточно быстро, чтобы это позволило объяснить даже почти мгновенное действие фитохрома. Синтез или расщепление гормона могли бы быть результатом прямого ферментативного действия Фдк или влияния Фдк на активность синтетических или гидролитических ферментов, существующих до облучения в виде "проферментов". Связывание или высвобождение гормонов в местах их функционирования или хранения могло бы происходить в результате модификации таких мест фитохромом Фдк.
Содержание четырех различных гормонов (гиббереллинов, цитокининов, этилена и ауксина) в тканях растений, выращенных в темноте, быстро изменяется после короткого облучения красным светом, тогда как уровень пятого (абсцизовой кислоты) изменяется при длительном влиянии красного света. Поскольку влияние красного света на гиббереллины, цитокинины и этилен можно снять дальним красным светом, фоторецептором для этих эффектов, несомненно, служит фитохром. Фдк повышает уровень гиббереллинов и цитокининов и снижает содержание ауксина и этилена. Изменения в содержании гиббереллинов наиболее детально изучены на этиопластах (пластиды из тканей, выращенных в темноте), выделенных из листьев ячменя. Краткое воздействие красным светом после выделения вызывает быстрый выход гиббереллина из пластиды и более медленное повышение уровня гормона, связанное, видимо, с его синтезом. Быстрые эффекты, вероятно, обусловлены изменением проницаемости мембраны этиопласта. У ячменя красный свет или гиббереллин вызывает раскручивание листьев.
Многочисленные другие быстрые эффекты превращения фитохрома тоже скорее всего связаны с изменениями в структуре и функции мембран. К этим эффектам относятся изменения в "притяжении" кончиков корней ячменя или маша к отрицательно заряженной поверхности (рис. 11.17), изменения в трансмембранном электрическом потенциале клеток и в "сонных движениях" листьев некоторых растений (рис. 11.18), которые в свою очередь зависят от изменений в передвижении ионов. Так как все эти процессы быстро реагируют на короткое облучение красным и дальним красным светом, они, несомненно, могут быть результатом изменений свойств мембран.
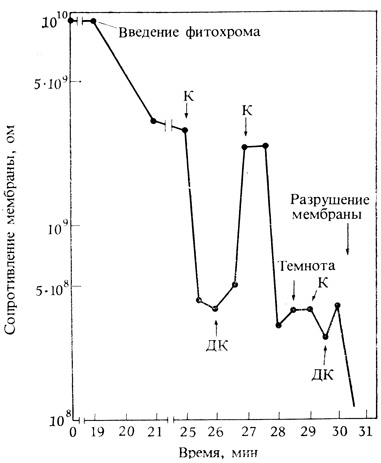
Рис. 11.19. Изменения в электрическом сопротивлении искусственной мембраны из окисленного холестерола при добавлении 6 мкг фитохрома в омывающий раствор и освещении красным и дальним красным светом. Обратите внимание, что красный свет снижает сопротивление, а дальний красный - повышает. (Roux, Yguerabide, 1973. Proc. Natl. Acad. Sci. 70, 702-764.)
Гипотеза о том, что фитохром действует на уровне мембран, подкрепляется также прямой демонстрацией включения фитохрома и его функционирования в искусственных липидных мембранах (см. гл. 2). Облучение таких мембран красным и дальним красным светом вызывает большие изменения в их. электрическом сопротивлении (рис 11.19). Это говорит в пользу того, что конформационные изменения хромофора фитохрома могут одновременно изменять и структуру мембраны. Другие, более медленные и отдаленные эффекты фитохрома, такие как активация генов и онтогенетические изменения, вероятно, могут быть следствием этого инициирующего события; но возможно, что фитохром сразу действует не в одном направлении. Как мы увидим в следующей главе, не все контролируемые фитохромом реакции локализованы в пределах одной клетки; в некоторых случаях облучение одной части растения влияет на развитие органов, находящихся на некотором расстоянии. Определенные реакции такого типа, контролируемые фитохромом, происходят в этиолированных тканях, но самые яркие примеры касаются фотопериодичеокой индукции цветения, клубнеобразования и покоя (см. гл. 12). Любая гипотеза о механизме действия фитохрома должна объяснять не только локальные, но и пространственно отдаленные реакции.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'