Движение воды в растении
Вода движется в растении по градиенту водного потенциала. Вода, поглощенная корневыми волосками и другими клетками эпидермиса, из клеток внешней части корня перемещается к ксилеме, занимающей центральную часть корня (рис. 6.8). Главным путем диффузии воды во внешней части корня служит апопласт - непрерывная совокупность клеточных стенок. Однако в эндодерме (цилиндрическом слое клеток, окружающем проводящую ткань) свободная диффузия по клеточным стенкам наталкивается на преграду - водонепроницаемый пробковый слой пояска Каспари. Вода должна изменить здесь свой путь и пройти сквозь мембрану и протопласт клеток эндодермы, играющей, таким образом, роль осмотического барьера между корой корня и его центральным цилиндром. У однодольных пробковеют также и внутренние тангенциальные стенки клеток, но эти стенки пронизаны порами, по которым, как по каналам, может проходить вода.
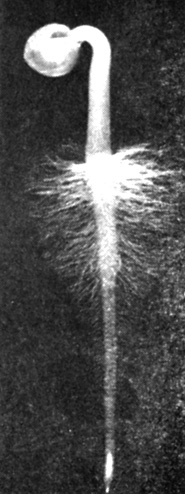
Рис. 6.7. Корешок проростка горчицы. Многочисленные тонкие выросты - корневые волоски
По ксилеме вода поднимается в надземные части растения. Ксилема состоит из нескольких типов клеток. Вода движется в ней главным образом по сосудам и трахеидам (рис. 2.6 и 6.9). И те и другие клетки прекрасно приспособлены для этой цели: они вытянуты в длину, лишены живого содержимого и внутри полые, т. е. это как бы трубки для воды. Одревесневшие вторичные клеточные стенки достаточно прочны на разрыв, чтобы выдерживать огромную разность давлений, возникающую при подъеме воды к вершинам высоких деревьев. Торцевые, а иногда и боковые стенки члеников сосудов перфорированы; сосуды, состоящие из соединенных конец в конец члеников, образуют длинные трубки, по которым легко проходит вода с растворенными в ней минеральными веществами. В трахеидах нет перфораций, и вода, для того чтобы попасть из одной трахеиды в другую, должна пройти через их торцевые стенки; однако трахеиды - очень длинные клетки, а потому и эта конструкция достаточно хорошо приспособлена для проведения воды. У цветковых растений есть и сосуды, и трахеиды; у более при-митивных форм сосудов, как правило, нет.
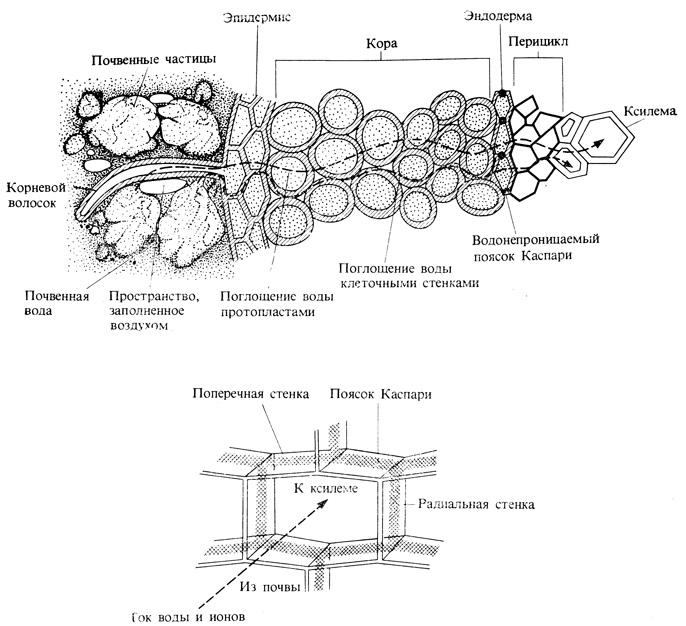
Рис. 6.8. Поступление воды из почвы в корень. Вода может перемещаться как по апопласту, так и по симпласту до тех пор, пока она не достигнет эндодермы. Дальнейшее передвижение по апопласту оказывается невозможным; здесь этот путь перекрыт барьером - пояском Каспари. В нижней части рисунка показан при большом увеличении поясок Каспари - водонепроницаемый барьер, заставляющий воду покинуть апопласт и устремиться через мембраны клеток эндодермы в симпласт
Листовые жилки, состоящие из тяжей ксилемы и флоэмы, образуют в листе настолько густую сеть, что любая его клетка оказывается достаточно близко от источника воды (рис. 3.9). Из ксилемы вода диффундирует в стенки клеток мезофилла. Таким образом, вода в жидкой фазе заполняет весь путь от почвы - через корень и стебель - до клеток мезофилла в листе. Суммарный поток воды направлен всегда в сторону меньшего водного потенциала, т. е. ψ максимален в почве, несколько ниже в клетках корня и самый низкий в клетках, примыкающих к эпидермимису листа. Малая величина ψ в этих последних клетках объясняется главным образом испарением воды с поверхности листа, т. е. транспирацией (которой посвящен следующий раздел). Табл. 6.2 дает представление о градиентах, обусловливающих движение воды в растении от ее поступления из почвы до испарения в атмосферу.
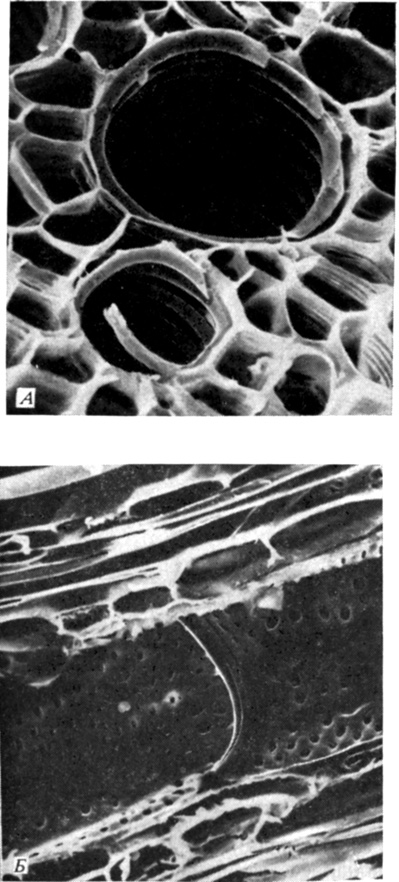
Рис. 6.9. А. Поперечный срез листового черешка клещевины (Ricinus communis) . Видны проводящие элементы ксилемы со спиральными утолщениями. Благодаря такому строению трубка может растягиваться по мере того, как черешок растет. (Диаметр более крупной трубки равен приблизительно 15 мкм.) Б. Продольный срез стебля Ricinus, на котором виден сосуд ксилемы и перфорированные торцевые стенки двух члеников сосуда в месте их соединения. Видны эллиптические и округлые окаймленные поры с замыкающей пленкой, но без торуса такие поры типичны для двудольных. (Диаметр сосуда равен приблизительно 21 мкм.) (С любезного разрешения Milburn J. A., Glasgow University; фотографии сделаны при помощи сканирующего электронного микроскопа.)
![Таблица 6.2 Примерные значения водного потенциала (ψ) и разности водных потенциалов (Δψ) для гипотетической системы почва - растение - воздух. [Предполагаемые условия - сравнительно небольшое дерево, хорошо увлажненная почва, относительная влажность воздуха около 50% при 22 °С (ψ=-1000 бар)1)]](pic/000132.jpg)
Таблица 6.2 Примерные значения водного потенциала (ψ) и разности водных потенциалов (Δψ) для гипотетической системы почва - растение - воздух. [Предполагаемые условия - сравнительно небольшое дерево, хорошо увлажненная почва, относительная влажность воздуха около 50% при 22 °С (ψ=-1000 бар)1)]
(1)R. G. S. Bidwell. 1974. Plant Physiology, New York, Macmillan.)
Вода переходит из растения в окружающий воздух главным образом в парообразном состоянии. В мезофилле листа имеются обширные межклеточные пространства, и каждая клетка мезофилла хотя бы одной своей стороной граничит с таким межклетником. Вследствие испарения воды с влажных клеточных стенок воздух в межклетниках насыщен водяными парами, и часть этих паров теряется - выходит наружу. Поскольку у большинства растений клетки эпидермиса покрыты воскообразной водонепроницаемой кутикулой, водяные пары выходят из листа в атмосферу главным образом через устьица (рис. 6.10).
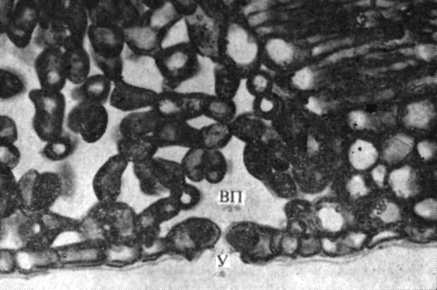
Рис. 6.10. Поперечный срез листа, на котором видно открытое устьице (У) с подустьичной воздушной полостью (ВП). Обратите внимание на то, что крупные, заполненные воздухом межклетники пронизывают всю толщу листа
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'