Синтез сахарозы и полисахаридов
В цикле Кальвина - Бенсона образуется, как уже отмечалось выше, фруктозо-6-фосфат (F-6-P). Этот гексозофосфат может под действием специфических ферментов превращаться в другие фосфорилированные гексозы, а именно в глюкозо-6-фосфат (G-6-P) и глюкозо-1-фосфат (G-1-P). Легко происходит и обратное превращение.
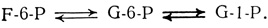
Из этих трех гексозофосфатов строятся затем цепи углеводных молекул, используемых для транспорта, хранения и в реакциях синтеза. Чтобы такие превращения могли произойти, гексозофосфаты предварительно должны быть активированы. Это обычно достигается в результате их присоединения к нуклеотидам - сложным кольцевым структурам, сходным с адениловой кислотой АТР. Продуктом такой реакции присоединения оказываются нуклеотидные производные моносахаридов, или нуклеотидсахара. Чаще других встречаются уридиндифосфоглюкоза (UDPG), образующаяся в реакции между уридинтрифосфатом (UTP) и глюкозо-1-фосфатом (G-1-P). Сам UTP образуется непрямым путем, в результате переноса фосфатной группы от АТР к UDP (уридиндифосфату).
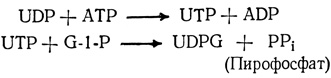
Нуклеотиды АТР и UTP присутствуют во всех клетках, потому что они используются наряду с другими нуклеотидами в синтезе ДНК и РНК.
Сахара транспортируются по растению в виде сахарозы - дисахарида, состоящего из остатков глюкозы и фруктозы (рис. 5.2). Сахароза образуется в реакции между UDPG и F-6-P:
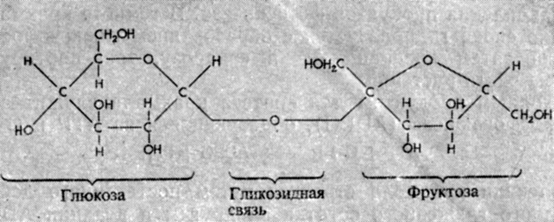
Рис. 5.2. Структура молекулы сахарозы (дисахарида)
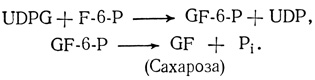
Равновесие этой реакции сильно сдвинуто в сторону синтеза сахарозы, чем обеспечивается возможность накопления данного дисахарида в значительных концентрациях. Для последующего использования сахароза должна предварительно подвергнуться расщеплению: фермент инвертаза катализирует ее гидролиз с образованием свободной глюкозы и фруктозы.
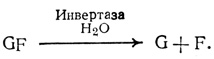
Энергия гликозидной связи в такой реакции растрачивается впустую, распределяясь между двумя молекулами. Поэтому если глюкозе и фруктозе предстоит распад в процессе дыхания или участие (в качестве сырья) в синтезе полисахаридов, то они должны предварительно вновь подвергнуться фосфорилирова- нию за счет АТР. Процессы синтеза и распада сахарозы наглядно показывают, что часто анаболические и катаболические реакции (реакции синтеза и распада) идут по разным путям.
Синтез крахмала и целлюлозы
Длинные полимерные цепи крахмала и целлюлозы построены из одних и тех же элементарных звеньев - остатков глюкозы, только соединенных по-разному. Это структурное различие обусловливает то, что два рассматриваемых полимера глюкозы (глюканы) существенно различаются по своей природе; крахмал, например, легко переваривается в организме человека" а целлюлоза совсем не переваривается. Главное же их различие состоит в том, что 1-й и 4-й углеродные атомы двух соседних остатков глюкозы соединены у крахмала α-связями, а у целлюлозы (β-связями (рис. 5.3). Крахмал представлен двумя формами: линейным полимером, или амилозой, не содержащим никаких других связей, кроме α-1,4-гликозидных, и разветвленным полимером, или амилопектином, в котором наряду с α-1,4-гликозидными связями имеются и 1,6-связи. Различие в характере связей определяет и неодинаковое пространственное расположение полимерных цепей. Крахмал - главный запасной полисахарид растения. Он нерастворим в воде и отлагается слой за слоем в крахмальных зернах, содержащихся в хлоропластах (см. рис. 2.20) или в лишенных хлорофилла лейкопластах запасающих тканей стебля, корней и семян. Иногда клетки запасающей ткани оказываются буквально забиты крахмальными зернами, которые легко в них выявить, поскольку они способны окрашиваться иодом в синий цвет. Будучи нерастворим в воде, крахмал в отличие от сахарозы и от гексоз не вызывает в клетках осмотического эффекта (см. гл. 6). Поэтому образование крахмала в клетках листа в периоды интенсивного фотосинтеза предотвращает подавление последнего, происходящее в результате накопления продуктов фотосинтеза. В темноте крахмал постепенно снова гидролизуется с образованием глюкозофосфата, который затем превращается в сахарозу, транспортируемую в другие части растения.
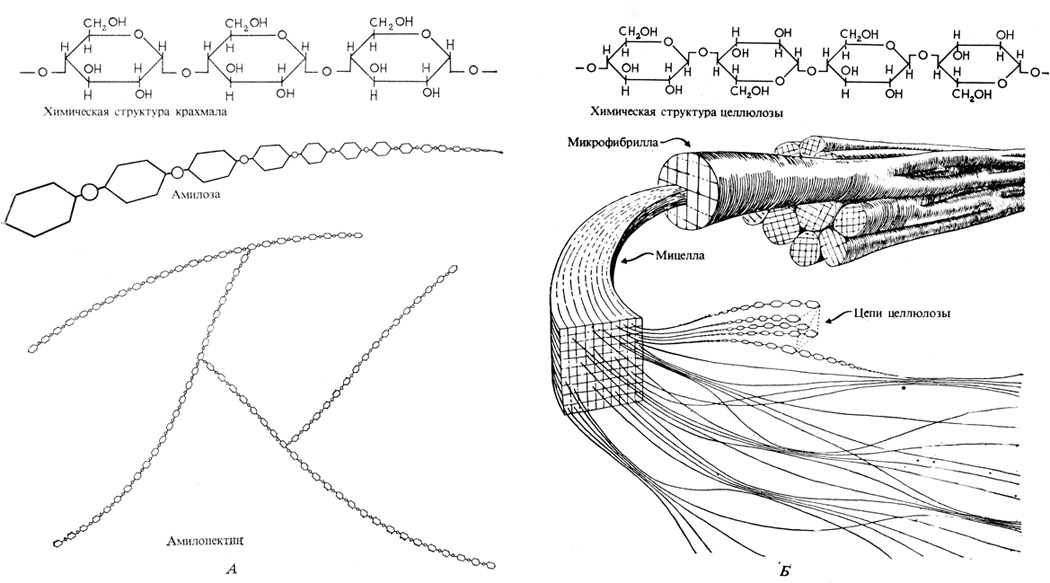
Рис. 5.3. Структура крахмала (А) и целлюлозы (Б) (С изменениями по J. Bonner, A. W. Galston. 1952. Principles of Plant Physiology, San Francisco, W. H. Freeman and Co.) Обратите внимание, что химические формулы крахмала и целлюлозы одинаковы, различаются же эти полисахариды пространственной ориентацией их кислородных мостиков. А. Крахмал, главный запасной полисахарид растения, построен из двух хорошо различимых компонентов: амилозы с ее длинными неразветвленными цепями из глюкозных звеньев и амилопектина, состоящего из большого числа коротких разветвленных цепей. Б. Целлюлоза, главный компонент первичной клеточной стенки, существует в виде длинных полимерных цепей. Цепи объединяются в мицеллярные тяжи, а последние - в микрофибриллы. Микрофибриллы, достаточно крупные для того, чтобы их можно было рассмотреть при помощи электронного микроскопа, составляют "основу" и "уток" клеточной стенки
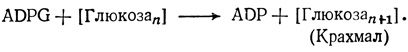
Исходным продуктом для синтеза крахмала служит аденозиндифосфоглюкоза (ADPG), образующаяся из АТР и G-1-P:

Молекула крахмала строится путем постепенного добавления одного глюкозного остатка за другим в реакции ADPG с предобразованной глюкозной цепью:
При низком содержании сахарозы крахмал расщепляется и. переводится в сахарозу. Однако сначала он расщепляется до остатков глюкозы и к каждому из них присоединяется остаток фосфорной кислоты, т. е. образуется G-1-P, чем обеспечивается сохранение энергии связи:
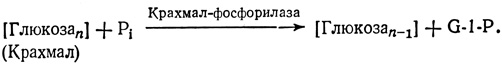
Этот G-1-P может затем использоваться для синтеза сахарозы, который мы описали выше. В семенах и в некоторых других органах, в которых одновременно идет расщепление больших количеств крахмала, он распадается до дисахарида мальтозы (G-G) под действием аα-амилазы. Мальтоза затем распадается до глюкозы, из которой (для транспорта) вновь синтезируется сахароза. На этом втором пути в отличие от первого энергия связи не сохраняется, поэтому здесь для превращения глюкозы в глюкозо-6-P требуется АТР.
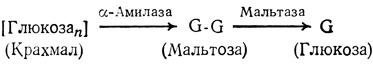
Целлюлоза, самый распространенный на Земле углевод, служит главным компонентом первичной клеточной стенки. Молекулы ее строятся подобно тому, как строятся молекулы крахмала, с тем, оцнако, отличием, что роль донора глюкозы играет другое нуклеотидное производное моносахарида - гуанозин- дифосфоглюкоза (GDPG) - и что связь между мономерными .звеньями принадлежит не к α-, а к β-типу.
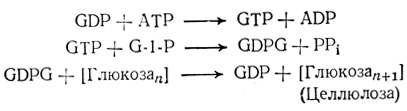
В некоторых случаях донором глюкозы для синтеза целлюлозы может быть и UDPG.
В организме высших растений целлюлоза расщепляется редко (если не считать распада, обусловленного деятельностью микробов). Два известных исключения из этого правила касаются клеток в отделительной зоне листа, образующейся перед сбрасыванием листьев, и сосудов ксилемы, у которых поперечные стенки растворяются. В отделительной зоне листа фермент целлюлаза разрушает клеточные стенки, расщепляя содержащуюся в них целлюлозу до отдельных мономерных единиц, т. е. до глюкозы. Клеточные стенки, ослабленные этим процессом, в конце концов разрываются, и лист сбрасывается.
Целлюлозные микрофибриллы в клеточной стенке скреплены при помощи матрикса из смешанных полисахаридных цепей, главным образом ксилоглюканов и арабиногалактанов (см. рис. 2.31). (Ксилоза и арабиноза - пятиуглеродные сахара (пентозы), а галактоза - гексоза, родственная глюкозе.) Эти полисахариды синтезируются также из предшественников, нуклео- тидсахаров, преимущественно в диктиосомах. Отшнуровывающиеся от диктиосом пузырьки в конце концов сливаются с плазмалеммой и таким путем передают свое содержимое формирующейся клеточной стенке.
Итак, все полисахариды легко переходят один в другой, но синтез их всегда идет через стадию нуклеотидсахаров, тогда как распад совершается более прямым путем.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'