Ареалы-области обитания растений
Видов растений - особенно семенных - на Земле великое множество, но распространены они по ее поверхности крайне неравномерно. Это сразу обнаруживается при сравнении числа видов, обитающих в разных районах планеты. Наиболее богаты видами тропики. Так, флора Индонезии насчитывает около 45 000 видов, а флора территорий, прилегающих к Амазонке, - по меньшей мере 40 000. По мере удаления от экваториальных областей число видов быстро уменьшается, и уже в субтропических пустынях оно очень невелико. На территории всей Сахары, например, встречается около 1200 видов. Но примыкающие к пустыням районы, занятые жестколистными лесами, снова обнаруживают большое видовое разнообразие: во флоре Средиземноморской области насчитывается по крайней мере 8000 видов, а в Калифорнии - примерно 6000. В умеренных широтах их число снова снижается - от более южных районов к более северным. Флоры таких стран, как Польша, ГДР, ФРГ или Чехословакия, где климатические условия примерно одинаковые, содержат 2200-3100 видов, Норвегии - 2100, во флоре Гренландии примерно 400, а на Шпицбергене - 130 видов семенных растений. В Антарктиде растут лишь два вида покрытосеменных.
Обратимся теперь к некоторым сведениям о распространении представителей семейства орхидных (Ochidaceae), которое уже давно привлекает внимание специалистов и ботаников-любителей. Оно содержит 20 000-25 000 видов и представляет собой самое богатое формами семейство среди всех покрытосеменных: ведь примерно каждый десятый вид цветковых растений - орхидея! Область наибольшего распространения орхидных, бесспорно, находится в тропиках, а именно в двух районах: в Южной и Юго-Восточной Азии (предполагают, что орхидные возникли именно здесь), где имеется не менее 6500-7000 видов, и в тропических районах Южной и Центральной Америки, где растет примерно 4500 видов. Что же касается Африки, то она значительно беднее орхидеями: здесь "только" около 2000 видов! К северу от тропиков число их быстро уменьшается. Во всем Средиземноморье едва наберется 150 видов (столько же и во всей Северной Америке), в Венгрии - 46, в Чехословакии - 53, в Польше - 49, в ГДР - 48 и в Скандинавии только 46 видов. Конечно, все эти сведения нельзя принимать как нечто абсолютное - в толковании видовых границ некоторых "критических" групп орхидей имеются значительные различия, а потому можно встретиться и с иными цифрами. Однако на общей тенденции это не отражается.
Величина и протяженность ареалов. Сказанное выше косвенным образом свидетельствует также и о том, что ботаники знают уже давно, а именно: определенные виды растений встречаются только на определенных территориях, иными словами, области их распространения ограниченны. Территорию, на которой обитает какой-либо вид или другая таксономическая группа (например, род или семейство*, называют ареалом.
* (Главные (основные) ранги таксономических групп, или таксонов, в восходящем порядке следующие: вид, род, семейство, порядок, класс, отдел.)
Размеры ареалов разных растений, то есть занимаемые ими площади, крайне разнообразны, Разумеется, чем крупнее таксон, тем больше и территория, которую он занимает. Так, ареал семейства всегда имеет большую протяженность, чем ареал рода, а тем более одного вида, относящихся к этому семейству. Но и ареалы разных видов также могут быть самыми разными.
Космополиты. Имеется целый ряд видов, которые распространены почти повсеместно, на большинстве континентов. Их принято называть космополитами. К числу космополитов в первую очередь относятся виды, которые легко могут расселяться на большие расстояния; так бывает со многими растениями, размножающимися спорами, и с микроорганизмами. Например, споры плесневых грибов встречаются почти повсюду. Другой вопрос - находят ли они всюду условия, подходящие для их развития. То же можно сказать и о многих бактериях. И все же как среди микроорганизмов, так и среди мхов и папоротников, споры которых также с легкостью переносятся на большие расстояния, не столь много космополитов, как можно было бы ожидать. Из мохообразных почти по всему миру распространены печеночный мох маршанция многообразная (Marchantia polymorpha) или встречающийся почти во всех водосточных желобах и в других влажных, богатых азотом местах листостебельный мох бриум серебристый (Bryum argenteum), а также другие виды этого рода.
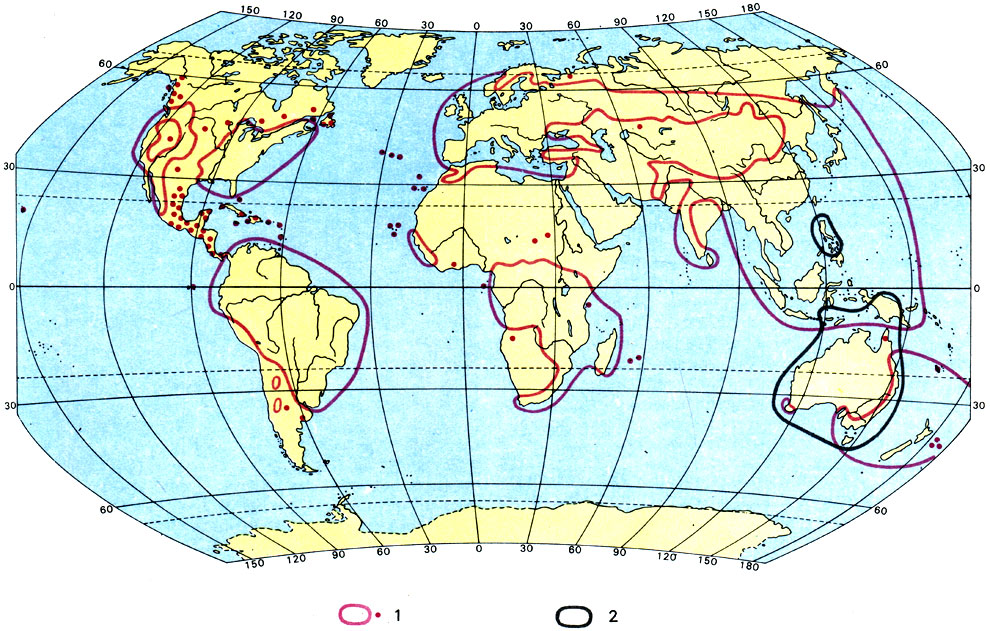
Распространение орляка обыкновенного (Pteridium aquilinum) (1) и видов эвкалипта (Eucalyptus) (2)
Среди папоротников классический пример космополитов представляет собой известный всем орляк обыкновенный (Pteridium aquilinum). Но каждый, кто интересуется окружающим его растительным миром, видимо, знает, что орляк встречается у нас отнюдь не повсюду. Кое-где можно обойти большие территории и не встретить ни одного экземпляра этого папоротника. Следовательно, космополиты - это не убиквисты, то есть растения, которые встречаются повсеместно и способны расти в самых разных местообитаниях. Орляк нуждается в определенных, пригодных для его жизни условиях. Прежде всего напомним, что растет он на кислых почвах. И хотя его требования к влажности почвы не очень жесткие, все же он избегает как сырых, так и слишком сухих почв. Строго говоря, истинных убиквистов среди высших растений нет. Каждый вид, даже если он очень широко распространен, предъявляет определенные требования к месту обитания. Остается сомнительным, существуют ли вообще такие условия местообитаний, которые распространены более или менее повсеместно.
К повсеместно распространенным видам относится целый ряд водных и болотных растений: тростник обыкновенный (Phragmites communis), частуха подорожниковая (Alisma plantago-aquatica), ряска малая (Lemna minor), виды рдеста (Potamogeton) и др. Несомненно, широкому расселению этих растений способствуют водоплавающие птицы, легко разносящие семена и части вегетативных органов (см. стр. 49), но и сама вода играет, вероятно, ведущую роль как главный фактор, нивелирующий условия местообитаний.
Если перечисленные выше виды широко распространены в природных местообитаниях и, следовательно, представляют собой примеры спонтанных космополитов, то у многих сорняков, а также у растений, живущих на замусоренных участках, и у придорожных растений обстоятельства, которые обусловили их космополитизм, существенно иные. Многие из этих растений ныне также распространены повсеместно, например марь белая (Chenopodium album), пастушья сумка (Capsella bursa-pastoris), крапивы двудомная и жгучая (Urtica dioica и Urtica urens), звездчатка средняя, или мокрица (Stellaria media), птичья гречиха (Polygonum aviculare), мятлик однолетний (Poa annua), подорожник большой (Plantago major) и др. Столь широким распространением эти виды обязаны человеку, это так называемые антропогенные космополиты. Их широкому расселению во многом способствовали мировая торговля и международные транспортные связи.
Говоря о космополитах, не следует забывать, что почти всегда речь идет об очень пластичных, богатых разными формами таксонах, включающих в себя множество разновидностей и т. п. Вероятно, это даже обусловливает возможность заселения ими столь обширных ареалов. По-видимому, лишь в редчайших случаях можно говорить о совершенно "единообразных" видах, как, например, о ряске малой (Lemna minor) и крапиве жгучей (Urtica urens), которые в этом отношении можно считать достойными внимания исключениями.
Эндемики. Полной противоположностью космополитам оказываются виды, которые произрастают только на какой-либо одной, очень ограниченной территории. Такие эндемичные виды часто встречаются лишь на каком-нибудь одном острове, в одной долине или на одном горном хребте. Так, например, только в пустыне Намиб на юге Африки растет вельвичия удивительная (Welwitschia mirabilis), занимающая особое место в системе растений.

Вельвичия удивительная (Welwitschia mirablis) - эндемик пустыни Намиб (Намибия)
Что касается величины ареала, то здесь понятие "эндемик" строго не ограничено. Те или иные таксоны, характерные для какого-либо континента в целом или для его части, также часто называют эндемичными. К их числу относятся род эвкалипт (Eucalyptus) или замечательные травяные деревья (род Xanthorrhoea), растущие в Австралии, а также южноафриканское семейство аизовых (Aizoaceae), включающее в себя суккулентные растения, и др. Эндемичными видами особенно богаты естественно ограниченные территории, на которых флора долгое время развивалась изолированно, - горные районы или острова. Последние тем богаче эндемиками, чем дальше они удалены от материков и чем дольше находились в изоляции. Современные Британские острова отделились от континентальной Европы около 7000 лет назад. Этот сравнительно небольшой срок, а также очень незначительное удаление от материка не позволили развиться там особым эндемичным видам. Напротив, во флорах средиземноморских островов Корсика и Сардиния, находившихся в изоляции более длительное время, обнаруживается примерно 5%, а во флоре острова Крит - около 10% эндемичных видов. Еще больше их на Канарских островах (36%), Мадагаскаре, отделившемся от Африки не менее 50 млн. лет назад (66%), или же на далеко отстоящих от материков Новой Зеландии (72%), Гавайских островах (82-90%) и острове Святой Елены (85%). На этих изолированных участках суши образовались не только эндемичные виды, но и эндемичные роды и даже семейства. Об островных эндемиках можно говорить как о формах, которые там возникли. Их называют прогрессивными эндемиками или неоэндемиками. То же можно сказать и о эндемичных таксонах горных областей.
Но имеются эндемичные виды, возникновение которых нельзя объяснить изоляцией. Так, например, выращиваемое в наших парках восточно- азиатское дерево гинкго (Ginkgo biloba) в настоящее время в диком виде растет только в провинции Чжэцзян в южном Китае. Однако ископаемые остатки свидетельствуют, что в прежние геологические периоды - юрском, меловом и третичном - род Ginkgo был представлен целым рядом видов, которые были распространены почти по всему северному полушарию. Таким образом, его современное местонахождение, бесспорно, оказывается только остатком некогда значительно большего ареала, и здесь эндемизм определяется тем, что Ginkgo - реликтовое растение.

Гинкго (Ginkgo biloba) - эндемик-реликт Восточной Азии -дерево, которое часто выращивают в парках
С таким же эндемизмом мы встречаемся и у метасеквойи (Metasequoia glyptostroboides). В 1944 г. произошло сенсационное для ботаников открытие этого вида хвойных деревьев, достигающих 30-метровой высоты, в горных лесах южнокитайской провинции Сычуань. Не говоря уже о том, что обнаружение в XX в. столь крупного представителя хвойных само по себе исключительная редкость, эта находка представляет особый интерес и потому, что метасеквойю считали вымершей миллионы лет назад. В отложениях третичного периода разных районов земного шара встречались остатки древесин, которые не принадлежали ни мамонтову дереву (Sequoia), ни болотному кипарису (Taxodium), а скорее занимали промежуточное положение между ними и поэтому были названы метасеквойей (слово meta означает "между"). Такие остатки найдены в верхнемеловых отложениях и встречаются в породах, которые образовались на протяжении всего третичного периода, то есть возникших за период времени почти 100 млн. лет. Остатки метасеквойи обнаружены по всему северному полушарию - на Аляске, в Северной Америке, в Гренландии, Шпицбергене, в Европе и от Сибири до Восточной Азии (рис. на стр. 18). И вот, как оказалось теперь, этот вид существует и поныне! Должно быть, это один из очень немногих случаев, когда растение сперва было известно как ископаемое, а затем уже было найдено как все еще существующее. Современный ареал Metasequoia крайне ограничен; этот вид - самый настоящий эндемик-реликт.
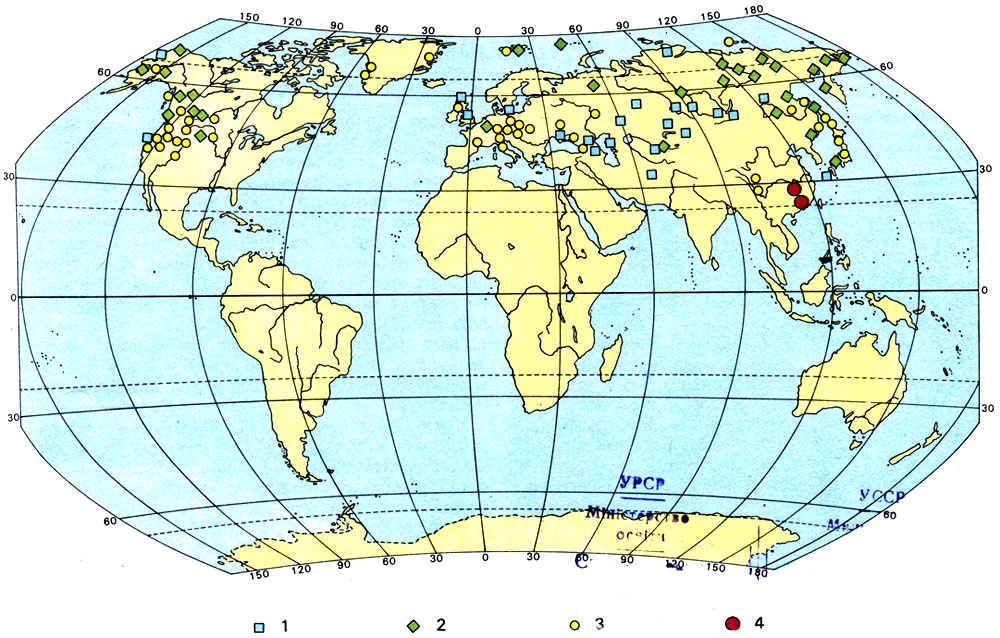
Распространение рода Ginkgo прежде и теперь: 1 - ископаемые остатки из юрских отложений; 2 - ископаемые остатки из меловых отложений; 3 - ископаемые остатки из третичных отложений; 4 - места современного произрастания в естественных условиях
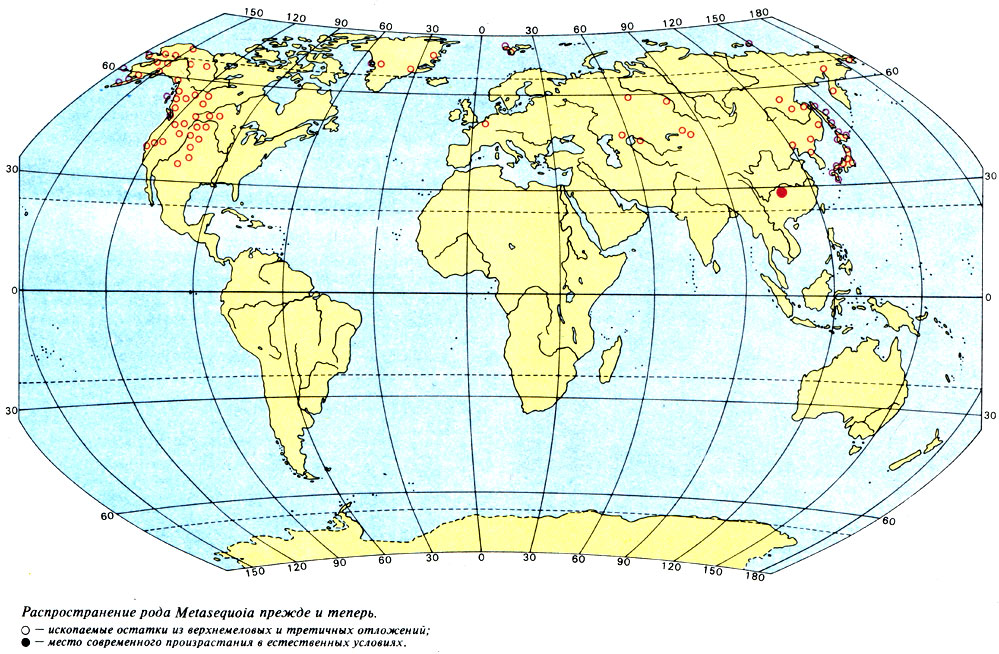
Распространение рода Metasequoia прежде и теперь. ○ - ископаемые остатки из верхнемеловых и третичных отложений; ● - место современного произрастания в естественных условиях
Мамонтовы деревья, или секвойи, два вида которых (гигантское мамонтово дерево, Sequoia gigantea, или Sequoiadendron giganteum, и вечнозеленое, Sequoia sempervirens) встречаются в некоторых районах на Западе США, - такие же эндемики-реликты. В третичном периоде они тоже были распространены по всему северному полушарию.
Число подобных примеров можно увеличить. Возникает вопрос: почему вид, ранее распространенный по разным континентам, в конце концов смог сохраниться на очень небольшой территории и стал эндемиком-реликтом? Ведь сам по себе термин ничего не говорит о причинах, обусловливающих возникновение такого эндемизма. Но, к сожалению, эти причины полностью не выяснены. Изменениями климата можно объяснить лишь вымирание вида в некоторых частях его ареала, например в Европе, поскольку неоднократное чередование холодных и теплых периодов в плейстоцене привело здесь к неоднократной же смене флор. А для полного восстановления исчезнувшей флоры предпосылок не существует.
Хотя уже упомянутая Welwitschia mirabilis, растущая в пустыне Намиб, благодаря ареалу, таксономической изолированности и другим признакам производит полное впечатление эндемика-реликта, все же это до сих пор не доказано. То же можно сказать и о многочисленных эндемиках Южной Африки (Капская флора), которые, с одной стороны, могут быть сочтены реликтами, а с другой - прогрессивными эндемиками.

Растущая в виде кустарника береза карликовая (Betula папа) - обычное тундровое растение, которое, однако, встречается и в Центральной Европе как ледниковый реликт
Даже в Центральной Европе, не имеющей четких естественных границ, встречаются эндемичные таксоны, но их немного. К их числу относятся растущий только у берегов Балтийского моря ситник балтийский (Juncus baltica), рябина шведская (Sorbus intermedia), распространение которой ограничено той же областью, и армерия боттендорфская (Armeria maritima subsp. bottendorfensis); последняя занимает крохотный ареал в южных предгорьях Гарца, где она растет на почвах, развившихся на медистых сланцах. Так же обстоит дело и с другими растениями, приспособившимися к обитанию на субстратах, содержащих тяжелые металлы. Но когда мы говорим о таких эндемиках, не ясно, идет ли речь о так называемых хороших (настоящих) видах, или лишь о возникших под влиянием внешней среды экологических расах одного вида (экотипах), или о чем-либо подобном. На страницах этой книги мы не будем подробно останавливаться на этом вопросе, но само его существование свидетельствует о том, что ареалогические исследования часто тесно связаны с систематикой. Достаточно сказать, что определение величины ареала во многих случаях зависит от того, в каком объеме понимается вид: чем шире это понятие, тем, естественно, крупнее ареал.

Первоцвет мучнистый (Primula farinosa), который изредка еще можно встретить на долинных болотах на севере ГДР; здесь это растение - ледниковый реликт
Поясним это на примере. Всем известная ветреница дубравная (Anemone nemorosa) встречается в Европе почти всюду, а также в Северной Америке и Восточной Азии и, следовательно, имеет весьма обширный ареал. Однако североамериканские растения некоторыми признаками отличаются от европейских, и их считают особым подвидом (subspecies quinquejolia). То же относится и к восточноазиатским представителям этого вида (subsp. amurensis). Если же объем вида понимать более узко и повысить ранг трех названных подвидов до ранга самостоятельных видов, то ареал Anemone nemorosa окажется находящимся только в пределах Европы.
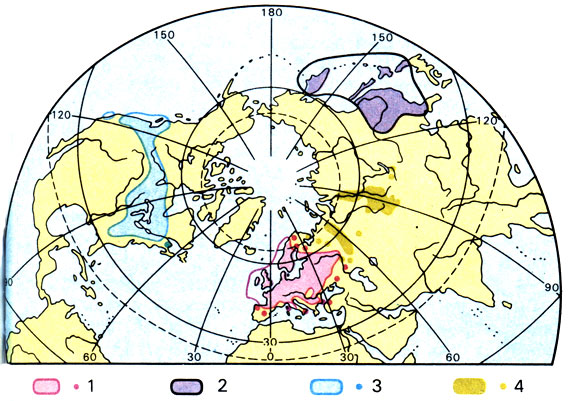
Распространение растений комплекса Anemone nemorosa (ветреница дубравная): 1 - Anemone nemorosa ssp. nemorosa; 2 - Anemone nemorosa ssp. amurensis; 3 - Anemone nemorosa ssp. quinquefolia; 4 - Anemone altaica
Викаризм. Приведенный пример свидетельствует о еще одном явлении, которое нередко наблюдается в мире растений: если какой-либо вид занимает большой ареал, то в некоторых частях последнего может происходить относительно самостоятельное развитие произрастающих растений, в результате чего начинается внутривидовая дифференциация. Такое развитие может способствовать не только возникновению разновидностей и подвидов, но и образованию особых видов. Именно так появляются ряды видов, географически в большей или меньшей мере замещающих друг друга. Близкородственные виды, сменяющие один другого, называют викарирующими, а это явление в целом - географическим викаризмом. Уточнение "географический" необходимо, поскольку имеется экологический викаризм, на котором мы остановимся ниже (стр. 42). Процессы такой дифференцировки происходят особенно быстро, если изначально единый ареал со временем разрывается.
Хорошим примером географического викаризма служат также виды лиственницы (Larix), которые в разных частях северного полушария образовали почти непрерывный ряд форм, замещающих одна другую (см. также т. 2).
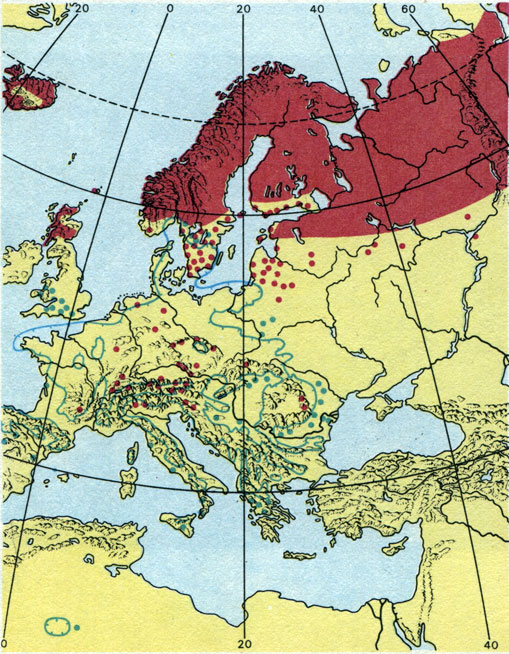
Распространение березы карликовой (Betula папа) в Европе
Сплошные и дизъюнктные ареалы, эксклавы и форпосты. Контуры ареалов столь же различны, как и их площади. Если растения того или иного вида произрастают на площади, ограничиваемой одной замкнутой линией, то говорят о сплошном ареале. Но часто представители какого-либо вида встречаются в нескольких изолированных районах. Если эти районы настолько удалены один от другого, что промежуточные пространства не могут быть преодолены с помощью естественных приспособлений для расселения, которыми обладают растения соответствующего вида, то речь идет о дизъюнктном, или разъединенном, ареале. С несомненными дизъюнкциями мы встречаемся, когда отдельные части ареала разъединены океаном или когда расстояния между ними достигают сотен и даже тысяч километров, как, например, в случае с ареалом упомянутой выше ветреницы дубравной. Разумеется, дизъюнкции бывают в ареалах не только видов, но также родов и семейств. Однако нередко все же не легко отличить сплошной ареал от дизъюнктного, так как обычно не известно, какие расстояния могут быть преодолены растениями какого-то вида с помощью свойственных им приспособлений для расселения (см. также стр. 33). Поэтому суждения о том, каков тот или иной ареал, часто бывают субъективными.
Многие растения по периферии своих ареалов встречаются относительно редко; здесь могут быть обнаружены так называемые эксклавы - небольшие участки ареалов, удаленные на сравнительно небольшие расстояния от основного, - или отдельные местообитания растений, встречающиеся вне основного замкнутого ареала. Такие местообитания называют форпостами; они могут либо свидетельствовать о происходящем расширении ареала, либо быть остатками некогда более обширного ареала. Часто форпосты и эксклавы имеют реликтовый характер. Таковы, например, местонахождения первоцвета мучнистого (Primula farinosa), встречающегося иногда на долинных болотах северо-восточного Мекленбурга, а также березы карликовой (Betula nana) в Гарце, Рудных горах и других горах Центральной Европы.
Итак, как мы имели возможность убедиться, области распространения растений нельзя считать постоянными: они все время изменяются, их пределы расширяются или сокращаются. Такие процессы происходили в течение всей истории развития растительного мира и происходят сейчас. Подробно разобраться в них не легко, для этого необходимо множество повсеместных наблюдений.
Знание ареалов растений представляет собой основную предпосылку для решения многих проблем геоботаники. Современный уровень знаний и еще далеко не полная изученность многих областей позволяют составить карты ареалов лишь небольшой части всех известных ныне видов, даже если при этом ограничиться только высшими растениями. Подробно закартированы лишь ареалы встречающихся в Европе растений (почти исключительно высших), и то далеко не всех. Большинство карт ареалов оказалось возможным создать только в самое последнее время. Таким образом, перед хорологией (так называют специальную область знаний, занимающуюся изучением распространения растений) стоит множество проблем.
Вряд ли нужно доказывать, что картирование ареалов не самоцель. Но карты помогают проводить дальнейшие исследования, такие, как сравнение и группировка (типизирование) ареалов, а прежде всего - их определение. Следующий этап - толкование ареалов, то есть выяснение вопроса о том, как возник тот или иной ареал, какие факторы определяли его становление и т. д. При этом прежде всего следует учитывать генетические, экологические, фитоценологические и исторические проблемы. На них мы остановимся в следующих разделах книги.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'