Физиологические группы анаэробных спороносных бактерий
Облигатные анаэробные бактерии, способные образовывать эндогенные споры, в настоящее время можно подразделить в зависимости от характера используемых ими субстратов и особенностей метаболизма на следующие физиологические группы: сахаролитические, протеолитические, пуринолитические и сульфатредуцирующие анаэробные бактерии.
Сахаролитические анаэробные бактерии способны сбраживать различные углеводы (органические кислоты и спирты, полисахариды, пектиновые вещества). Обычно конечными продуктами брожений являются органические кислоты, растворители (спирт, ацетон, бутанол и др.), углекислый газ и водород. Было установлено существование нескольких типов брожения.
Маслянокислое брожение осуществляют такие анаэробы, как CI. butyricum, CI. tyrobutyricum, CI. lactoacetophilum, и другие виды. Главные продукты брожения - масляная и уксусная кислоты. Так, сбраживание глюкозы GL butyricum можно описать уравнением
Образование масляной кислоты происходит также, если в среду вместо углеводов вносить молочную кислоту вместе с уксусной. Уксусная кислота играет роль акцептора водорода. Образовавшиеся при маслянокислом брожении кислоты далее микроорганизмом не потребляются.
Ацетонобутиловое брожение представляет собой биохимически более сложный тип маслянокислого брожения, в котором образовавшиеся на первой стадии жирные кислоты превращаются в нейтральные конечные продукты - ацетон, бутиловый и изопропиловый спирты. Если в первой фазе брожения увеличивается кислотность, то во второй фазе кислотность не только не возрастает, но даже уменьшается. Таким образом, за счет превращения кислот в нейтральные продукты регулируется рН среды (кислотность). Ацетонобутиловое брожение углеводов вызывается CI. acetobutylicum и близкими к нему по физиологии видами. CI. butylicum осуществляет брожение, очень сходное по типу с ацетонобутиловым, но отличается тем, что вместо ацетона в этом случае образуется изопропиловый спирт.
Уксуснокислое брожение углеводов наблюдается у CI. aceticum и CI. thermoaceticum. Уксусная кислота - единственный продукт брожения.
Пропионовокислое брожение присуще О. propionicum. В качестве основных продуктов образуются пропионовая и уксусная кислоты, а также углекислый газ.
Брожение пектиновых веществ (полиурониды) способны осуществлять анаэробные бактерии из различных систематических групп. В настоящее время известно около 18 видов клостридиев, способных разрушать пектиновые вещества. Наиболее активные пектинолити-ческие виды - GI. felsineum, CI. laniganii, GI. pectinolyticum, CI. pectinovorum, CI. virens и другие пигментированные и непигментированные клостридии и плектридии. Имеются и факультативно-анаэробные формы - CI. macerans и CI. polymyxa. Каждому из этих видов свойственны свои специфические детали обмена веществ, но общим их свойством является способность разлагать пектиновые вещества до органических кислот, спиртов и газов. Некоторые бактерии обладают очень стойкими пектинолитическими свойствами и выделяют пектинолитические ферменты на средах без пектиновых веществ. У других анаэробов (например, у CI. multifermentans) синтез ферментов происходит только при добавлении в среды пектинов (индуцированный синтез ферментов).
Сбраживание Сахаров пектинолитические анаэробы осуществляют по маслянокислому или ацетонобутиловому типу.
Брожение целлюлозы. Большая группа высокоспециализированных анаэробных спорообразующих бактерий получает энергию за счет сбраживания целлюлозы. Конечными продуктами брожения являются уксусная, пропионовая, масляная и молочная кислоты, этиловый спирт, водород и углекислый газ. В качестве промежуточных продуктов при гидролизе целлюлозы образуются глюкоза и целлобиоза. И весьма удивительно, что если добавить к среде глюкозу (или другие сахара), то они очень плохо усваиваются либо совсем не используются. А если в среде имеются одновременно глюкоза и клетчатка, то сбраживается прежде всего клетчатка. Это свидетельствует о высокой специализации целлюлозоразлагающих анаэробов.
Целлюлозолитические бактерии отличаются не только физиологическими, но и морфологическими особенностями. Большинство целлюлозолитических спорообразующих анаэробов имеют вид очень тонких длинных палочек, образующих споры по плектридиальному типу.
Обычно вегетативные клетки находятся в адсорбированном состоянии на волокнах целлюлозы (рис. 51). Вероятно, это связано с тем что ферменты, гидролизующие клетчатку (целлюлазы), в среду не выделяются, а прикреплены к поверхности клеток. Под микроскопом можно увидеть, что разрушаются те участки клетчатки, к которым клетки непосредственно прикасаются. Спорообразующие клетки обычно находятся в растворе, так как при спорообразовании изменяется характер связи со средой, а спорообразование идет за счет эндогенного метаболизма (за счет внутриклеточных запасов питательных веществ).
Первая культура анаэробных спорообразующих бактерий, способная осуществлять брожение целлюлозы, была выделена В. Л. Омелянским. В литературе описано 12 видов целлюлозолитических анаэробов. Спорообразующие целлюлозолитические анаэробы до настоящего времени очень плохо изучены в связи с трудностями их выделения и культивирования. Большинство выделенных штаммов были загрязнены посторонними бактериями, что значительно обесценивает данные по физиологии и биохимии этих организмов.
Брожение органических кислот и спиртов. Выделено несколько специализированных видов анаэробных бактерий (возможно, их в природе существует намного больше), использующих органические кислоты и спирты в качестве источника углерода и энергии.
Так, CI. kluyveri получает энергию путем сопряженного окисления-восстановления системы этиловый спирт - уксусная кислота. В результате реакции образуются высшие жирные кислоты (главным образом капроновая и масляная кислоты). Характерно, что CI. kluyveri не способен сбраживать углеводы, аминокислоты и пурины. Накопление энергии в форме АТФ происходит у этого анаэроба с использованием механизмов окислительного фосфорилирования. CI. lactoacetophilum осуществляет брожение сходного типа, сбраживая смесь лактата и уксусной кислоты. Обе эти бактерии используют в качестве акцепторов водорода уксусную кислоту, а в качестве доноров первая - этанол, вторая - лактат.
CI. glycollicum превращает этиленгликоль в смесь этилового спирта и уксусной кислоты:
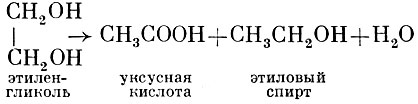
Анаэробная бактерия из рода Clostridium (не определенная до вида) осуществляет крайне интересный тип брожения соединений с сульфониевой связью. Такие вещества могут рассматриваться как соединения, обладающие макроэргической связью, при разрыве которой выделяется большое количество энергии. Clostridium species (Штадтман, 1968) сбраживает диметилпропиотетин, используя его в качестве единственного источника углерода и энергии:
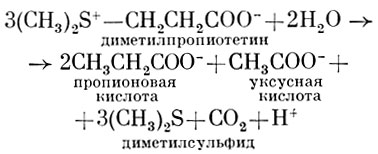
Протеолитические анаэробные бактерии - очень большая группа анаэробов, получающая энергию за счет анаэробного разрушения аминокислот. Многие из них являются строго протеолитическими организмами, неспособными сбраживать углеводы. Другие обладают слабовыраженной сахаролитической активностью. Имеются также активные протеолитические виды, способные осуществлять маслянокислое брожение Сахаров. Протеолитическая активность у некоторых анаэробов (CI. histolyticum) настолько высока, что у зараженных этими микроорганизмами животных наблюдается как бы расплавление мышц, после чего остаются голые кости. Освобождающиеся после протеолиза (разрушение белков протеолитическими ферментами) аминокислоты могут сбраживаться двумя путями. CI. sporogenes, например, сбраживает пару аминокислот, причем одна из них окисляется, а другая восстанавливается (реакция Стикленда), т. е. одна аминокислота служит донором, а другая - акцептором водорода:
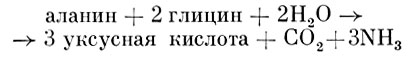
Существуют виды протеолитических анаэробов, которые могут сбраживать и по одной аминокислоте, например глутаминовую кислоту (CI. tetani).
Пуринолитические анаэробы. Существуют три вида бактерий, сбраживающих гетероциклические соединения с азотным кольцом: СI. acidiurici, CI. cylindrosporum и CI. uracilicum. Они способны разрушать пурины и пиримидины с образованием уксусной кислоты, углекислого газа и аммиака. Первые два вида не способны использовать ни углеводы, ни белки (аминокислоты). Ими быстро расщепляются ксантин, гуанин, гуанозин, 6,8-диоксипурин; более медленно и после адаптации (приспособления) разрушаются гипоксантин и инозин. CI. acidiurici и CI. cylindrosporum физиологически очень близки, но четко разделяются по морфологическим признакам. У CI. acidiurici споры имеют овальную форму и расположены терминально. Клетки при спорообразовании раздуваются. У CI. cylindrosporum споры цилиндрические, расположены центрально или субтерминально. Спорангии при этом не раздуваются.
Сульфатредуцирующие бактерии. Восстановление микроорганизмами полностью или частично окисленных соединений минеральной серы в анаэробных условиях за счет окисления органического вещества или водорода называют микробной сульфатредукцией. Этот процесс был открыт русским химиком Н. Д. Зелинским в 1890 - 1893 гг. Чистая культура сульфатредуцирующих бактерий была выделена Бейеринком в 1895 г. Сначала бактерий, восстанавливающих сульфаты, относили к неспороносным. Затем было замечено, что в культурах сульфатредуцирующих бактерий иногда появляются споры (когда инкубирование проводилось при высоких температурах). В настоящее время выяснилось, что большинство неспорообразующих штаммов, выделенных из различных субстратов, было загрязнено спорообразующими. Последние выделены в новый род Desulfotomaculum, в который входят четыре вида: D. nigrificans, D. ruminis, D. orientis, D. antarcticum. Первые два вида используют некоторые органические кислоты (молочную, пировиноградную), но не усваивают углеводы (глюкозу). D. antarcticum способен усваивать глюкозу. D. nigrificans - термофил, оптимальная температура для роста этого вида 55° С. Восстановление сульфатов происходит по уравнению:
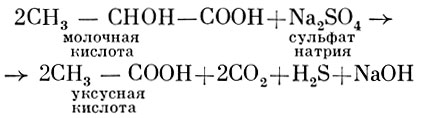
Таким образом, у анаэробных спорообразую-щих бактерий мы встречаемся с удивительным физиологическим разнообразием как по механизмам анаэробного получения энергии, так и по диапазону используемых субстратов.
Имеется группа анаэробов, использующих главным образом углеводы; другая группа разлагает преимущественно белки и аминокислоты; третья - сбраживает только пурины; четвертая - получает энергию за счет окисления органических кислот или водорода, используя в качестве акцептора водорода сульфаты.
Специфичность по отношению к субстратам у спорообразующих анаэробов выражена резко. Потребности в питательных веществах у этих групп весьма различны. Некоторые протеолитические анаэробы (например, GI. sporogens) не удовлетворяются средами, содержащими полный набор аминокислот, углеводы, минеральные соли, комплекс витаминов, активаторы микробного роста. Такие резко выраженные гетеротрофы растут лишь на средах, содержащих белки или продукты их частичного гидролиза. С другой стороны, существуют анаэробы, развивающиеся на очень простых средах, в состав которых входит несколько минеральных солей, в том числе сульфаты и органическая кислота либо водород в качестве донора электронов (сульфатредуцирующие бактерии). При этом может усваиваться атмосферный азот.
Способность фиксировать молекулярный азот широко распространена среди спороносных бактерий. Этот процесс могут осуществлять маслянокислые, ацетонобутиловые и сульфатредуцирующие бактерии. Наиболее активные азотфиксаторы - сахаролитические анаэробы (клостридии).
Отношение к кислороду не одинаковое у различных физиологических групп спорообразующих анаэробов. Более устойчивыми к действию кислорода являются сахаролитические анаэробы. Часть представителей этой группы - аэротолерантные формы (способные давать слабый рост на пластинках агара даже в аэробных условиях), например CI. carnis, CI. histolyticum и др. Очень чувствительны к кислороду и трудно культивируются сульфатредуцирующие бактерии. Они могут развиваться только в строго анаэробных условиях при тщательном удалении кислорода из сред, в которых они культивируются.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'