Ископаемые хвощевидные
Самые древние достоверные ископаемые остатки хвощевидных известны из верхнедевонских отложений. При этом уже в позднем девоне существовали представители всех классов отдела. Столь далеко зашедшая к концу девона дифференциация в пределах хвощевидных сама по себе свидетельствует о древности этой группы. Если принять во внимание сложную организацию некоторых позднедевонских представителей отдела (например, стробилов у эвиостахии - Eviostachya hoegii (рис. 76), то придется признать, что, хотя более древние его представители и не обнаружены, в целом хвощевидные к концу девона прошли столь же длительный путь эволюции, как и плауновидные.
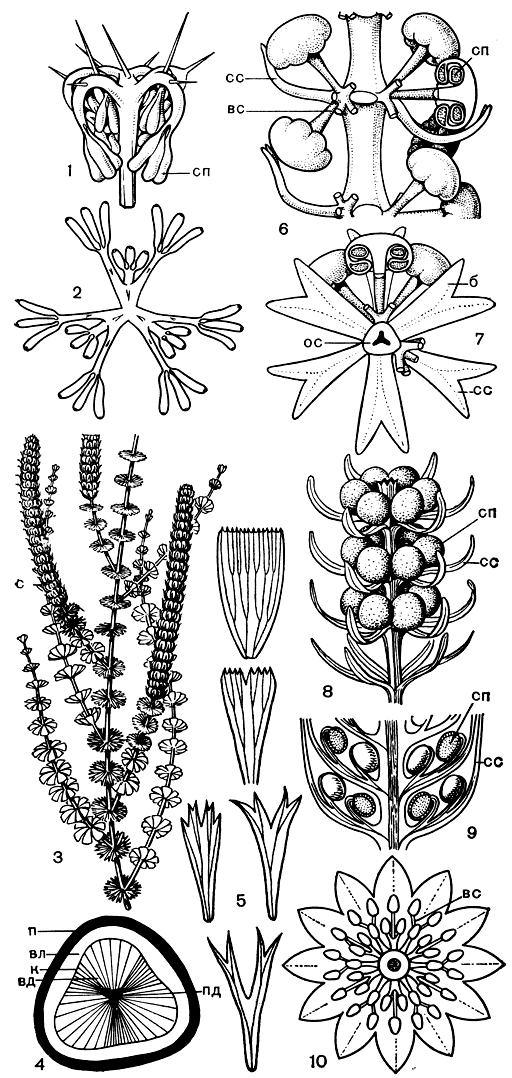
Рис. 76. Клинолистовые (Sphenophyllopsida): 1, 2 - эвиостахия Хёга (Eviostachya hoegii) (1 - спорангиофор в прижизненном положении, 2 - спорангиофор в развернутом виде); 3-5 - клинолист клинолистный (Sphenophyllum cuneifolium) (3 - общий вид части растения, 4 - схема поперечного сечения стебля в области междоузлия, 5 - листья разных узлов стебля); 6-10 - стробилы представителей порядка клинолистовых: 6-7 - пельтастробус Рид (Peltastrobus reedae) (б - часть стробила сбоку после удаления брактей и некоторых веточек спороносных сегментов спорангиофоров, 7 - поперечный срез стробила в области междоузлия - некоторые веточки спороносных сегментов удалены), 8 - сфенофиллостахис водяной (Sphenophyllostachys aquensis); 9-10 - сфенофиллостахис Досона (S. dawsonii): 9 - продольный срез части стробила, 10 - поперечный срез стробила в области междоузлия; (сп - спорангий; к - камбий; вд - вторичная древесина; вл - вторичный луб; п - перидерма; пд - первичная древесина; cm - стробил; б - брактея; вс - веточка спороносного сегмента спорангиофора; ос - ось стробила; сс - стерильный сегмент спорангиофора)
Членистость стеблей - характерный признак хвощевидных - неясно выражена лишь у представителей класса гиениопсид, небольшого размера растений, существовавших в среднем девоне (роды гиения - Hyenia и каламофит - Calamophyton). Дихотомическое ветвление стебля гиениопсид еще больше удаляет их от двух других классов хвощевидных - клинолистовых (Sphenophyllopsida) и собственно хвощовых (Equisetopsida). Листовые органы гиениопсид трехмерно дихотомически ветвятся. Их веточная природа проявляется, таким образом, чрезвычайно отчетливо.
Споронесущие структуры гиениопсид представляют настоящую загадку для морфологов: спорангии расположены по одному (каламофит) или по три (гиения) на концах дихотомически разветвленных осей, относительно природы которых у ботаников нет единодушного мнения. Одни называют их спорофиллами (признавая за ними листовое происхождение); другие - спорангиофорами (считая, что применение этого термина не обязывает к суждению о происхождении органа). Спорангиеносный орган гиениопсид является как бы неусложненным еще спорангиофором современных хвощей - с меньшим количеством спорангиев и пока лишенный явно защитного образования - щитка (именно щитки - шестиугольные плотно сомкнутые образования - бросаются нам в глаза при разглядывании незрелых стробилов хвощей). Кроме того, эти "неоформившиеся спорангиофоры" гиениевых расположены рыхло на оси - плотная защитная упаковка еще не выработалась.
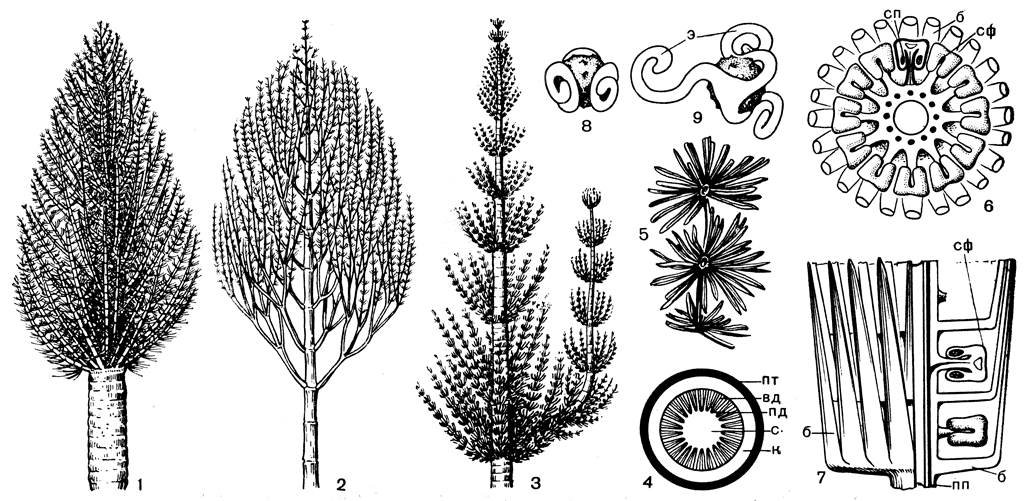
Рис. 77. Ископаемые хвощовые (Equisetopsida): 1-3 - каламиты (Calamites) с разными типами ветвления; 4 - схема поперечного сечения междоузлия стебля каламита: вд - вторичная древесина, к - кора, пд - первичная древесина, пт - покровная ткань, с - сердцевина, 5 - листья каламитов; 6 - поперечный срез стробила каламита (Galamostachys) в области междоузлия; 7 - часть стробила, вид сбоку: слева - с поверхности, справа - в продольном разрезе; (б - брактея, пп - проводящий пучок, сп - спорангий, сф - спорангиофор; 8 - спора элатеритеса (Elaterites triferens) во влажном состоянии; 9 - то же - в сухом состоянии, э - элатера)
Если дихотомическое ветвление всех структур и конечное расположение спорангиев (признаки, буквально "лежащие на поверхности") сближали гиениопсиды с риниофитами и зостерофиллофитами, то анатомическое строение их стебля, исследованное в последние годы, указывает на иное родство - с одним из вымерших классов папоротниковидных - кладоксилопсидами (Cladoxylopsida), существовавшими в раннем девоне.
Клинолисты (род Sphenophyllum, рис. 76), образовывавшие густые заросли в лесах каменноугольного периода, изучены лучше других вымерших хвощевидных. При жизни побеги некоторых растений, по-видимому, превышали в длину несколько метров. Поскольку диаметр даже обладавших вторичным приростом стеблей незначителен (0,1-0,5 см, редко до 1,5 см), можно думать, что наряду с полегающими стеблями у клинолистов были и лазящие стебли; об этом могут свидетельствовать и прицепки, обнаруженные на стеблях одного из видов.
К узлам более молодых участков стебля прикреплялись мутовки обычно из 6-9 клиновидных листьев. Многим, если не всем, клинолистам была свойственна гетерофиллия (разнолистность).
При относительном однообразии своего внешнего облика многочисленные представители клинолистовых резко различались между собой по строению стробилов, располагавшихся на концах ветвей (рис. 76).
Расцвет класса хвощовых также приходится на каменноугольный период. Древесный ярус заболоченных тропических лесов в это время в значительной мере состоял из каламитов (Calamites). По облику каламиты несколько напоминают хвощи, но только увеличенные в десятки раз (рис. 77). Обладавшие центральной полостью и довольно мощным (до 12 см) слоем вторичной древесины стволы каламитов нередко превышали в диаметре 0,5 м, достигая в высоту 8-10 м (редко 20 м). Основными фотосинтезирующими органами каламитов были расположенные мутовчато листья (рис. 77), длина которых у разных видов варьировала от 2 мм до 7 см. Число и размеры членов мутовки изменялись в пределах одного растения в зависимости от относительного возраста узла. В мутовке могло быть от 3 до 70 листьев.
Разнообразные по строению стробилы каламитов достигали иногда в длину 12 см и в диаметре 4 см. Стробилы были смешанными, они состояли из чередующихся мутовок стерильных листьев и спорангиофоров (рис. 77). Не исключено, что все каламиты были разноспоровыми растениями, производящими обычно однополые стробилы. У одного из каламитов найден мегастробил (каламокарпон замечательный - Calamocarpon insignis), в спорангии которого развивалась всего одна мегаспора. Эта спора прорастала внутри спорангия, образуя женский гаметофит. Такая крайняя степень редукции числа мегаспор в спорангии отмечена и у древовидных плауновидных палеозоя.
У каламитов найдены также стробилы, в спорангиях которых сохранились споры с элатерами (рис. 77). Исследователь этих спор американский палеоботаник Р. Бакстер сообщил в своей статье, что Споры вымершего около 300 млн. лет назад элатеритеса (Elaterites), к его удивлению, вдруг зашевелились под микроскопом, повинуясь быстрым движениям своих элатер! Это произошло в тот момент, когда Бакстер переносил споры из водной среды в спиртовую. По-видимому, иссушающее действие спирта произвело на них тот же эффект, который вызывается и подсыханием на воздухе спор хвоща. В финале этого удивительного зрелища, наблюдаемого Бакстером, ископаемые элатеры в результате своих собственных движений отламывались и плавали в препарате, как таинственные вопросительные знаки.
К концу палеозоя - началу мезозоя каламитовые, достигшие, вероятно, крайней степени специализации, при резком изменении климата Земли вымерли. Менее специализированные родичи каламитовых - хвощовые (Equisetaceae) пережили эпоху резкого изменения климата и в первой половине мезозоя в некоторых областях Земли даже образовывали почти чистые сообщества по берегам водоемов. В конце концов почти все представители хвощевидных также вымерли, и от некогда процветавшего на Земле отдела до наших дней дошел лишь род хвощ.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'