Семейство раффлезиевые (Rafflesiaceae)
Семейство раффлезиевых включает 8 родов и не более 55 видов, распространенных главным образом в тропических странах. Роды раффлезиевых группируются в 4 трибы, или, по Р. Торну (1976), подсемейства.
Подсемейство цитиновых (Cytinoideae) включает два рода - цитинус (Cytinus) и бдаллофитон (Bdallophyton). В роде цитинус 6 видов, распространенных в Средиземноморской области, в Малой Азии, на Черноморском побережье Кавказа (в районе Пицундской сосновой рощи), в Южной Африке и на Мадагаскаре. Род бдаллофитон содержит 4 вида, распространенных в Мексике и Сальвадоре. Цитиновые паразитируют на различных представителях семейства ладаныиковых.
В подсемейство аподаытовых (Apodanthoideae) входят роды аподантес (Apodanthes) и пилостилес (Pilostyles). Род аподантес, насчитывающий 5 видов, распространен в тропической Америке, где паразитирует на представителях семейства флакуртиевых. Род пилостилес (около 28 видов) распространен главным образом в Америке (от южных штатов США до Магелланова пролива), но 2 вида встречаются в Южной Африке, 1 вид - в Западной Австралии и 1 вид - в Турции, Ираке и Иране. Виды пилостилеса паразитируют на кустарниках из семейства бобовых, а единственный азиатский вид - пилостилес Гаусскнехта (P. hausskneehtii) - паразитирует на некоторых видах астрагалов, главным образом из подрода трагаканта (подрод Tragacantha рода Astragalus), а однажды был найден на чингиле (Halimodendron halodendron), также относящемся к семейству бобовых.
Подсемейство раффлезиевых (Rafflesioideae) состоит из трех родов - раффлезия (Rafflesia), саприя (Sapria) и ризантес (Rhizanthes). Все три рода распространены в тропических лесах Азии и на прилегающих островах и паразитируют на корнях лиан из семейства Виноградовых, особенно на корнях циссуса (Cissus) и тетрастигмы (Tetrastigma). В роде раффлезия около 12 видов, произрастающих в Западной Малезии (полуостров Малакка, острова Суматра, Ява, Калимантан, Филиппины). Саприя (вероятно, только 1 вид) распространена в Северо-Восточной Индии и в Индокитае. Ареал рода ризантес (1 - 2 вида) приблизительно совпадает с ареалом раффлезии (полуостров Малакка, острова Суматра, Ява, Калимантан).
Наконец, подсемейство митрастемоновых (Mitrastemonoideae) образует род митрастемоп (Mitrastemon), включающий 5 видов, которые распространены в Южной Японии и на острове Тайвань, в Индокитае, на островах Суматра и Калимантан, в Южной Мексике и Гватемале (2 вида). Митрастемон паразитирует на корнях представителей семейства буковых.
Раффлезиевые изучены к настоящему времени еще далеко не полно и неравномерно. Это обусловлено их обитанием в труднодоступных или малонаселенных областях, а также их биологией. Большую часть жизни они проводят в тканях растений-хозяев или в почве. Тем не менее многие особенности их биологии и развития уже стали достоянием науки. Проростки раффлезиевых внедряются обычно в корни растения-хозяина посредством присосок (гаусториев). Те части проростка, которые остаются вне тканей растения-хозяина, постепенно отмирают, и дальнейшее развитие паразита происходит интра-матрикально, т. е. исключительно в тканях корня или стебля растения-жертвы. Есть основания считать, что прорастание и внедрение паразита происходит здесь теми же способами, что и у других столь же высокоспециализированных, но значительно лучше изученных паразитных растений (например, из семейства заразиховых). Об этом говорят сходные особенности строения их семян и высокая степень избирательности растений-хозяев. Можно предполагать, таким образом, что, так же как и в семействе заразиховых, семена раффлезие-вых побуждаются к прорастанию выделениями из корней растений-хозяев, эти же выделения ориентируют направление роста проростков, а внедрение гаусториев в ткани растений-хозяев происходит в результате действия специальных ферментов, разрушающих их клетки. Все развитие происходит в дальнейшем, как мы уже отметили, интраматрикально. Обитающее в тканях растения-хозяина тело паразита называют эндофитом, а такой способ паразитизма - эндопаразитизмом. Эндофит получает питание из окружающих клеток растения-хозяина.
В отношении большинства родов довольно определенно известно, что их проростки внедряются в корни растений-хозяев. Некоторые исследователи считают, однако, что, в отличие от них, растения из родов пилостилес и аподантес паразитируют исключительно на стеблях. Тем не менее прямые наблюдения прорастания этих паразитов на ветвях растений-хозяев отсутствуют. В то же время особенности строения семян пилостилеса и аподантеса и особенности их распространения свидетельствуют о том, что их проростки внедряются, равно как и проростки других раффлезиевых, вероятно, сначала в корни растений-хозяев и лишь затем врастают в ткани стеблевых частей растения. О том, что такой переход, видимо, возможен, свидетельствует пример некоторых видов цитинуса, для которых определенно известно внедрение проростка в ткани корня; в то же время цветки некоторых видов этого рода нередко появляются на поверхности стеблей растений-хозяев. На стеблях лиан наблюдались даже цветки крупноцветковых видов раффлезии. Не исключено, однако, что отдельные, случайно попавшие на ветки семена паразита могут здесь прорасти и внедриться.
В тканях растений-хозяев тело паразита распространяется в виде клеточных тяжей, напоминающих грибные гифы. Их расположение, строение, ветвление и другие особенности развития могут быть довольно разнообразны. Иногда они окружают (как у аподантеса и цитинуса) центральную часть стебля или корня растения-хозяина довольно плотным сетчатым чехлом, о чем можно судить по распределению цветков или плодущих побегов паразита (рис. 85).
Цветки раффлезиевых закладываются эндогенно. В некоторых участках растения-паразита происходит образование зачатков отдельных цветков (раффлезия и др.) или побегов, на которых впоследствии образуется целое соцветие (цитинус и бдаллофитон). Эти зачатки растут, развиваются и, наконец, появляются наружу через разрывы покровных тканей растения-хозяина. На более ранних стадиях развития зачатки цветков или побегов растения-паразита можно определить по характерным бугоркооб-разным вздутиям корней или стеблей растения-жертвы.
У раффлезии одиночные цветки появляются на поверхности пораженного корня растения-хозяина в последовательном порядке, то реже, то чаще. У видов, эндофит которых обычно проникает в стебли растений-хозяев, происходит почти одновременное и кучное появление бутонов, которые распускаются тут же, прилегая к коре растения-хозяина (виды пилостилеса). У видов, корни растений-хозяев которых расположены сравнительно глубоко в почве и эндофит которых не способен проникнуть в стебель, на поверхности корня появляются мясистые побеги, имеющие вместо зеленых листьев лишь довольно твердые, кожистые защитные чешуи. Так, у средиземноморского цитинуса красного (Суtinus rubra, рис. 85), встречающегося также в Пицундской роще, побеги паразита довольно дружно появляются на поверхности корней ладанника (Cistus) без определенного порядка в зависимости от того, насколько сильно поражен корень и насколько хорошо эндофитный паразит снабжается питательными веществами (табл. 16); Побеги, зацветающие этой весной, закладывались и появлялись на поверхности корней летом предыдущего года. Здесь же на корне видны и следы отмерших прошлогодних побегов. Довольно часто зачатки побегов цитинуса красного появляются столь кучно, что создается впечатление одного ветвящегося побега.
Для пилостилеса Гаусскнехта отмечена интересная зависимость расположения его цветков на стебле от положения листьев растения-хозяина. Листья растения-хозяина и цветки паразита возникают на стебле синхронно, с равной периодичностью. Это происходит потому, что клетки эндофита проникают в точку роста стебля растения-хозяина и здесь происходит общая регуляция развития листьев астрагала и цветков паразита. Такое явление ботаники называют изофазным паразитизмом. Изофазный эндопаразитизм - высшая степень приспособления цветковых растений паразитов к чужеядному существованию.
Цветки раффлезиевых от очень мелких до необычайно крупных, одиночные и сидячие или реже в колосовидных соцветиях (большинство видов цитинуса и бдаллофитона), большей частью однополые, редко полигамные или обоеполые. Большинство видов семейства раффлезиевых имеют двудомные цветки. Однодомные цветки характерны, например, для цитинуса красного. Обоеполые цветки известны у митрастемона и ризантеса, но у последнего встречаются, кроме того, и мужские цветки, т. е. цветки его полигамные. Полигамные цветки наблюдаются также у бдаллофитона и отдельных видов раффлезии. В однополых цветках встречаются рудименты цветков другого пола. Цветки раффлезиевых безлепестные. Чашелистиков обычно 4 - 5, сросшихся в нижней части в трубку или редко свободных, черепитчатых или редко створчатых (ризантес), иногда лепестковидных. Тычинок от 5 до многих, более или менее сросшихся в трубку или чаще в мясистую колонку. Пыльники в 1 - 3 рядах вокруг трубки или колонки, вскрывающиеся продольной или поперечной щелью или верхушечной порой. Пыльцевые зерна 3 - 4-бороздные, многобороздные, 2 - 3-поровые или безапертурные. Гинецей состоит обычно из 4 - 10 (у митрастемона 9 - 20) плодолистиков; рыльце сидячее или на столбике, дисковидное, головчатое или многолопастное. Завязь нижняя или полунижняя, редко верхняя (митрастемон), одногнездная или ложномногогнездная (раффлезиевые), с очень многочисленными анатропными (аподантовые и митрастемон) или чаще ортотропными семязачатками, расположенными на впяченных внутрь полости завязи (интрузивных) планцентах или покрывающих всю стенку завязи. Семязачатки битегмальные (аподантовые) или чаще унитегмальные. Плоды ягодообразные. Семена многочисленные, мелкие, с недифференцированным зародышем и маслянистым эндоспермом,
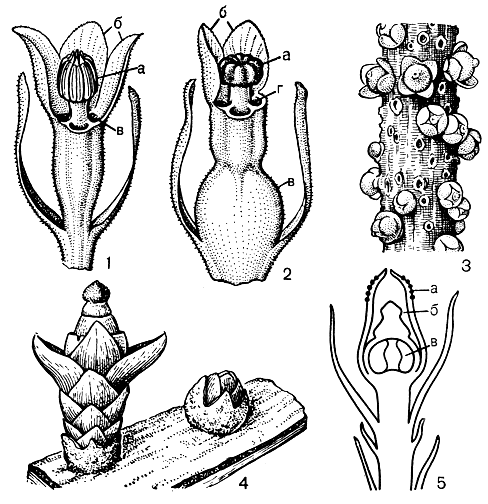
Рис. 85. Цветки некоторых представителей семейства раффлезиевых: 1 - мужской цветок цитинуса красного (Cytinus rubra) (а - андроцей, б - околоцветник, в - каналы, ведущие к нектарникам); 2 - женский цветок цитинуса красного (а - рыльце, б - околоцветник, в - завязь, г - каналы, ведущие к нектарникам); 3 - цветки аподантеса (Apodanthes caseariae) на стебле растения-хозяина; 4 - бутон и цветок митрастемона ямамоты (Mitrastemon jamamotoi) на стебле растения-хозяина; 5 - схема строения цветка митрастемона (а - синандрий, б - рыльце, в - полость завязи с долями плаценты)
В подсемейство раффлезиевых входят наиболее известные представители этого семейства. Цветки у них всегда крупные, с мясистым околоцветником. В некоторых случаях они достигают действительно рекордной величины (до 1 м в поперечнике, как у раффлезии туан-мудэ - Rafflesia tuan-mudae, и около 45 см у раффлезии Арнольда - R. arnoldii, табл. 15). Тем не менее и самые маленькие из них выглядят вполне внушительно. Так, у яванской раффлезии патма (R. patma) цветки часто достигают в диаметре 30 см, у саприи - 20 см, у ризантеса - 10 - 15 см. Цветки этих растений выделяются, однако, не только размерами, но и строением, окраской, специфическим запахом.
Гигантские цветки раффлезии привлекают внимание насекомых-опылителей тем, что напоминают и видом и запахом разлагающиеся куски мяса (табл. 15). На запах и вид гниющего мяса слетаются целые рои мух. Основная окраска листочков околоцветника раффлезии от кирпично-красной до темно-коричневой, иногда с пурпурным оттенком. Отмеченный выше эффект гниющего мяса подчеркивается светлыми, неправильной формы, нерегулярно расположенными пятнами, которые выделяются на общей поверхности.
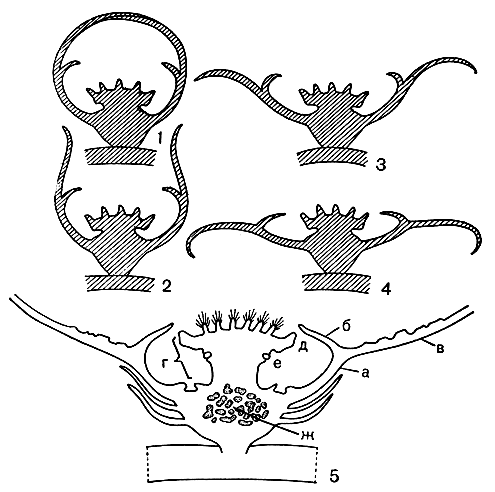
Рис. 86. Развитие и строение цветка раффлезии (Rafflesia): 1 - 4 - последовательные этапы раскрытия бутона; 5 - схема строения цветка (а - трубка околоцветника, б - диафрагма, в - краевая доля околоцветника, г - колонка, д - диск, е - зона расположения пыльников, ж - доли плаценты в полости завязи)
Общий план строения обоеполого цветка раффлезии можно видеть на рисунке 86. В строении цветков раффлезии привлекает внимание прежде всего массивная колонка в центре цветка над завязью, где в одно целое соединены андроцей и гинецей. Кроме того, интересно особое разрастание околоцветника, образующее так называемую диафрагму, характерную только для цветков раффлезии и саприи. Диафрагма окрашена светлее в сравнении с околоцветником, что, вероятно, имеет некоторое значение при опылении. Вершина колонки разрастается в стороны, и эта расширенная ее часть называется диском. Ниже его краев вокруг колонки располагаются пыльники. У раффлезии они погружены в отделенные друг от друга углубления. У родственных родов саприя и ризантес пыльники ясно двухгнездные, у раффлезии каждый пыльник состоит из нескольких гнезд. Все они раскрываются через верхушечные поры. Пыльцевые зерна 3 - 4-бороздные. Важная особенность пыльцевых зерен раффлезии и ризантеса состоит в том, что зрелая пыльца здесь не одиночная, а собирается в отдельные, соединенные слизистым веществом комочки. Эта слизь, вероятнее всего, образуется из дегенерировавших остатков стенок пыльцевых гнезд.
Нижняя завязь раффлезии ложномногогнездная. Такая завязь, так же как у ризантеса и саприи, образуется в результате разрастания серии париетальных плацент, закладывающихся в виде пластинок и впоследствии образующих многочисленные срастания. На поперечных срезах через завязь (рис. 87) можно видеть последовательные этапы эволюции плацентации в семействе. Важно отметить при этом, что подобные разрастания плацентарной ткани характерны именно для представителей этого подсемейства крупноцветковых видов и связаны с особенностями распространения семян.
Хотя раффлезия пользовалась особым вниманием исследователей, точные данные о биологии опыления ее цветков неизвестны, неизвестно даже, например, где на диске располагается воспринимающая пыльцу ткань. У раффлезии туанмудэ наблюдали прорастание пыльцы на внешних краях диска, но не исключено, что и шиповидные выросты на его поверхности также могут способствовать улавливанию и прорастанию пыльцевых зерен.
Развитие раффлезии и ее огромных цветков происходит очень медленно. В Богорском ботаническом саду (Индонезия) для выяснения этого вопроса ставили специальные опыты, и оказалось, что от высева семян до появления бутонов над поверхностью почвы прошло три года и еще полтора года понадобилось для того, чтобы показавшийся над почвой бутон превратился в открытый цветок. Тем более удивительно, что время функционирования цветков раффлезии очень коротко, всего 2 - 4 суток. Еще более удивительно то, что раффлезия цветет далеко не часто.
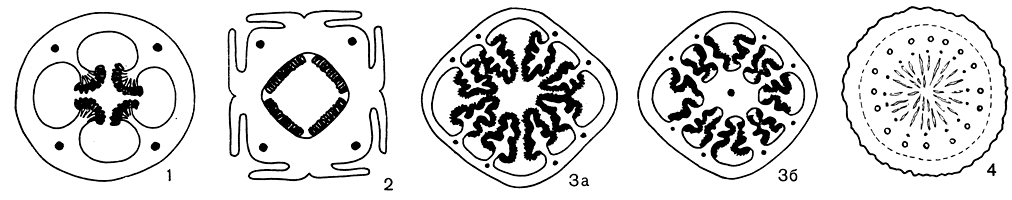
Рис. 87. Строение завязи у некоторых представителей семейства раффлезиевых. Усложнение плацентации в процессе исторического развития: 1 - поперечный медианный разрез завязи аподантеса Прингля (Apodanthes pringlei); 2 - поперечный медианный разрез завязи аподантеса флакуртиевого (A. flacourtiae); 3 - поперечный разрез через апикальную (а) и базальную (б) части завязи цитинуса красного (Cytinus rubra); 4 - поперечный медианный разрез завязи раффлезии патма (Rafflesia patma)
Объединение пыльцы в слизистые комочки, окраска цветков и их специфический запах указывают на то, что цветки раффлезии и других крупноцветковых родов опыляются насекомыми, предпочитающими гнилостные продукты, вероятно разными видами лесных мух. Ягодообразные плоды содержат вязкую массу (пульпу), в которую погружены многочисленные семена. Это приспособление для эпизоохорного распространения семян раффлезии. Считается, что ее семена разносят дикие свиньи и слоны, к конечностям которых прилипают части раздавленных плодов, содержащих множество семян. Цветки раффлезии обычно находят в местах, где почва обильно удобрена экскрементами слонов. Возможно, что мелкие млекопитающие и насекомые тоже способствуют дальнейшему распространению семян в окружающей местности, где часть из них оказывается в непосредственной близости от корней растений-хозяев, распространяющихся в самом поверхностном слое почвы. Своеобразная форма семян с их придатками типа элайосом, возможно, служит приманкой для муравьев. Раффлезия - вероятно, единственное известное растение, семена которого могут распространяться как слонами, так и муравьями.
В подсемействе аподантовых, включающем роды пилостилес и аподантес, мы находим, напротив, одни из самых мелких цветков. Тем не менее принципиальное их устройство в основном такое же, как и у гигантских цветков раффлезии. Цветки, по-видимому, всегда однополые. В мужских цветках пилостилеса Турбера (Pilostyles thurberi) тычинки овальные, пыльники, раскрывающиеся поперечной щелью, располагаются в три ряда вокруг вершины центральной колонки. У пилостилеса Гольца (P. holtzii) центральная колонка является собственно рудиментом гинецея, а тычинки срослись и образуют вокруг колонки тычиночную трубку, увенчанную многочисленными пыльниками. Семязачатки у пилостилеса располагаются обычно по всей внутренней поверхности стенки завязи. У аподантеса можно различить четыре плацентарных выроста, усеянных семязачатками (рис. 87). Переходные формы встречаются у пилостилеса. В отличие от подсемейства раффлезиевых анатропные семязачатки аподантовых битегмальные. О механизмах опыления ничего не известно. Известно только, что цветки невзрачных, буроватых оттенков, а пыльцевые зерна одиночные.
Зародыши пилостилеса еще более редуцированы, чем зародыши видов подсемейства раффлезиевых, строение же семян и их величина в общем сходные, хотя и имеются некоторые различия в деталях строения.
О распространении семян пилостилеса и аподантеса почти ничего не известно. Некоторые исследователи предполагают, что крысы, муравьи и даже термиты разносят плоды и семена этих растений и вносят их в почву. Не исключена и роль ветра и дождевых вод. Если семена прорастают прямо на ветвях растений-хозяев, то в распространении семян могут участвовать и птицы. Но особенности строения семян определенно указывают на их прорастание в почве и внедрение в корни растений-хозяев.
Довольно своеобразно устроены цветки митрастемона - единственного представителя митрастемоновых (рис. 85). Одиночные обоеполые цветки венчают вершину небольшого побега. Тычиночная трубка имеет большое число пыльников, раскрывающихся продольной щелью. Особенность этой тычиночной трубки состоит в том, что она окружает гинецей и возвышается над ним, оставляя на вершине лишь небольшое отверстие. Это приспособление тесно связано с особенностями опыления. Цветок протандричен, и во время созревания пыльников насекомые не могут проникнуть через узкое отверстие на вершине тычиночной трубки внутрь цветка к завязи. Так предотвращается самоопыление. Несколько позже гинецей, вырастая, разрушает тычиночную трубку, и в это время поверхность рыльца способна принять пыльцу с других растений.
Предполагают, что опыление цветков митрастемона происходит при посредстве мух, в частности плодовых мушек (Drosophila). He исключено и участие в опылении птиц из рода белоглазка (Zosterops). Во всяком случае, отмечено накопление больших количеств нектара в чашеподобных верхних кроющих листьях-чешуях. Семязачатки расположены в завязи на разрастающихся к ее центру пластинчатых париетальных плацентах. Зародыш крайне редуцирован и состоит всего из четырех клеток. Общее строение семени такое же, как и у других раффлезиевых, но семена еще мельче. Способы распространения семян неизвестны.
Подсемейство цитиновых - единственное в семействе, у большинства представителей которого цветки собраны в соцветия. Весьма оригинальны соцветия у бдаллофитона. У одних видов цитинуса, например у цитинуса двудомного (Cytinus dioicus), цветки двудомные, в то время как у других, например у цитинуса красного, они однодомные. Продолговатые пыльники в мужских цветках цитинуса собраны на вершине колонки в так называвмый синандрий. У бдаллофитона каждый пыльник увенчан оригинальной шпорой, то менее, то более выраженной у разных видов. Пыльники двухгнездные и раскрываются продольными щелями. У обоих родов воспринимающая поверхность головчатого рыльца радиально-выемчатая, образующая овальные доли. Многочисленные семязачатки цитинуса расположены на радиальных плацентах. Цветки цитинуса красного опыляются пчелами. Детали механизма опыления неизвестны.
Так же как и у раффлезии, в основании семязачатка цитинуса образуются разрастания (элайосомы), указывающие на распространение его семян припочвенными животными.
Раффлезиевые не имеют практического значения, но представляют интереснейший объект для научных исследований.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'