Порядок Зеленые, или Настоящие, мхи - Bryales
Этот порядок занимает в типе мохообразных центральное положение. По некоторым структурным признакам, например по анатомическому строению стебля, зеленые мхи стоят ближе к сосудистым растениям, чем другие мохообразные.
От порядка сфагновых зеленые мхи отличаются отсутствием водоносных клеток в листьях и на поверхности стебля, наличием многоклеточных ризоидов и более сложным строением спорогона, коробочка которого сидит обычно на длинной ножке.
По условиям местообитаний и по образу жизни мхи порядка Bryales более разнородны, чем сфагновые мхи. Зеленые мхи широко распространены в лесах, в особенности хвойных, в тундре, в горах, на лугах и в степи; некоторые виды обитают в проточной воде рек и ручьев, а также на камнях, стволах деревьев и т. п. С таким широким экологическим диапазоном зеленых мхов связано и их морфологическое разнообразие: порядок Bryales охватывает 620 родов с общим числом видов до 13 000.
Кукушкин лен (Polytrichum commune) -самый крупный из зеленых мхов; он широко распространен в сыроватых лесах и на болотах, где обитает рядом со сфагнумом, занимая относительно высокие участки микрорельефа. Почти чистые заросли кукушкина льна образуют в лесах высокие, пышные темно-зеленые подушки.
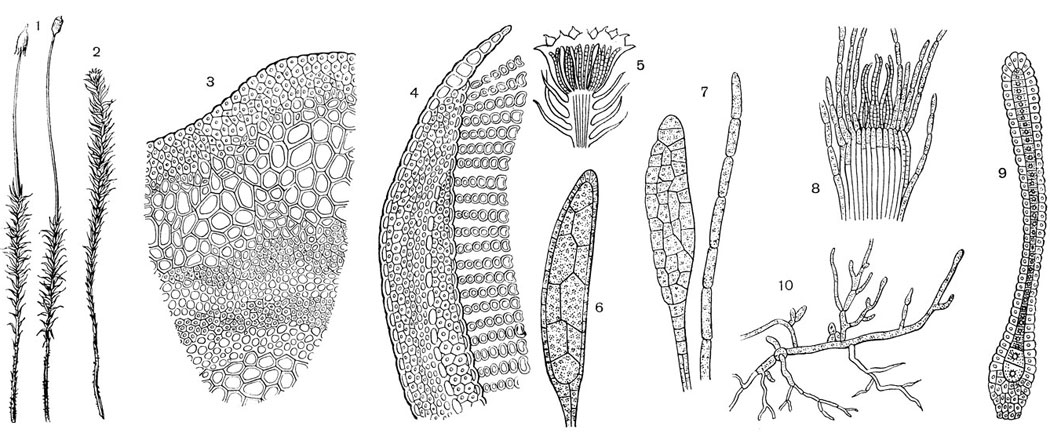
Рис. 53. Кукушкин лен (Polytrichum commune): 1 - женское растение со спорогоном; 2 - мужское растение с розеткой на верхушке; 3 - поперечный разрез стебля; в центре рисунка - листовой след; 4 - поперечный разрез листа; справа видны ассимиляторы; 5 - верхушка мужского растения с антеридиями; 6 - антеридий: ножка, стенка, спермагенная ткань; 7 - парафизы; 5 - верхушка женского растения с архегониями; 9 - архегоний: брюшко; с яйцеклеткой, шейка с канальцевыми клетками; 10 - протонема с почками
Строение вегетативных органов кукушкина льна лучше изучать на живых растениях, которые содержат в лаборатории в сходных условиях с описанными для содержания сфагнума (стр. 119). Можно использовать для занятий и сухой кукушкин лен, размоченный в горячей воде перед занятиями.
Верхушки мужских и женских растений, содержащие органы полового размножения, заготовляют весной и хранят в спирте. Спорогоны с колпачками собирают позже - в конце весны или начале лета, хранят также в разбавленном спирте. Развитие споро-гона кукушкина льна, начинаясь во второй половине лета - осенью, прерывается зимой и заканчивается только в начале лета следующего года.
Внешнее строение (рис. 53, 1, 2). Длинные, прямые и довольно крепкие стебли кукушкина льна густо покрыты сравнительно длинными, жестковатыми листьями. Более старые листья становятся бурыми и, отмирая, отваливаются. На подземной безлистной части стебля, называемой часто "корневищем", находятся ризоиды, образующие слегка скрученные пучки.
В отличие от одноклеточных ризоидов печеночников ризоиды зеленых мхов представляют однорядные, многоклеточные, тонкие, бесцветные нити.
Рассмотрим в лупу листостебельное растение. Отделим иглами от стебля несколько листьев, состоящих из линейной остроконечной пластинки с острыми шипиками по краям и широкого пленчатого влагалища, почти обхватывающего стебель. Листья располагаются тесной спиралью, влагалища их находят одно на другое так, что стебель покрывается ими со всех сторон, как футляром.
Внутреннее строение листа (рис. 53, 3). Стебель с прижатыми к нему листьями зажимают в сердцевину бузины и делают серию срезов. Изготовив препарат, изучают его при малом и большом увеличении микроскопа.
Анатомическое строение листовой пластинки кукушкина льна своеобразно. На нижней поверхности листа находится эпидермис; на краях лист тонок и однослоен, а в толстой многослойной середине его находится проводящий пучок (53, 4). Ассимиляционная ткань лежит открыто на верхней поверхности листа. В поперечном срезе она представляется ярко-зелеными вертикально стоящими колонками, каждая из которых состоит из одного ряда хлорофил-лоносных клеток. Эти колонки выносятся на одинаковую высоту и делают весь срез похожим на гребенку. Легко представить, что на листе кукушкина льна находятся продольные и довольно высокие пластинки-ассимиляторы. Между тесно стоящими пластинками хорошо удерживается вода, всасываемая зелеными клетками. В сухую погоду края листа, завертываясь кверху, смыкаются над ассимиляционными пластинками, защищая их от высыхания.
Размножение. Половое воспроизведение, размножение и весь цикл развития зеленых мхов совершаются по общему плану развития мохообразных. Поэтому ограничимся лишь изучением некоторых особенностей строения антеридиев и архегониев, их размещением на растениях, а также строением спорогона.
Побеги с антеридиями отличаются по розетковидным верхушкам. Листочки розетки окрашены в красновато-желтый цвет, они более широкие и короткие в сравнении со стеблевыми листьями. В центре розетки, на верхушке стебля, между парафизами сидят антеридии на толстых многоклеточных ножках (рис. 53, 5).
При сборе материала в мае - начале июня зрелость антеридиев устанавливается таким образом. Если слегка сдавить пальцами розетку, то из нее выступает капля мутно-белой жидкости, состоящей из ослизнившихся материнских клеток сперматозоидов; если сжать розетку сильнее, то выдавливаются белые продолговатые тельца: это и есть антеридии. Из заспиртованных розеток антеридии также легко выделяются. Крупные антеридии имеют вытянутую и слегка изогнутую форму, они заполнены густым содержимым. Кроме антеридиев, в препарате обычны довольно разнообразные, прозрачные парафизы. Надавив осторожно иглой на покровное стекло, добиваемся выхода из верхушек антеридиев массы спермагенных клеток. При этом обнаруживается, что стенка антеридия состоит из одного слоя прозенхимных клеток. В естественных условиях клетки на верхушке зрелого антеридия разъединяются и материнские клетки сперматозоидов выходят наружу (рис. 53, 6, 7).
Мужские растения, после опадения антеридиев, продолжают рост, оставляя на стебле листочки розетки. По числу розеток можно определить, сколько лет данный побег производил антеридии.
Найти и рассмотреть архегонии кукушкина льна несколько труднее, так как верхушки женских побегов, где находятся архегонии, ничем не отличаются от вегетативных побегов. Чтобы выделить группу архегониев, надо сначала осторожно раздвинуть и удалить листья, окружающие их. Архегонии на одной верхушке находятся в разных стадиях развития; зрелые - отличаются от архегониев маршанции более длинной шейкой и большим количеством канальцевых клеток, а также массивной ножкой (рис. 53, 8, 9). .
В природе перемещение сперматозоидов с мужских растений на женские происходит при тех же обстоятельствах, как у маршанции (стр. 115). Половой процесс у кукушкина льна доступен для наблюдения. Методика работы описана на стр. 115.
Зрелый спорогон кукушкина льна состоит из коробочки, прикрытой волокнистым колпачком, и длинной ножки, основание которой, глубоко внедряясь в верхушку женского растения, является гаусторией (рис. 54). Коробочка спорогона состоит из урны и крышечки с заостренной верхушкой.
Первое время после оплодотворения брюшко архегония и развивающийся спорогон растут равномерно; затем, вследствие ускорения роста спорогона, стенка брюшка разрывается. Верхняя часть брюшка с остатками шейки архегония выносится вверх растущим спорогоном и остается в виде защитного колпачка на его коробочке и в конце концов сдувается ветром. Нижняя часть брюшка продолжает расти вместе с верхушкой стебля, образуя длинное влагалище, плотно прилегающее к гаустории спорогона. Позже, когда спорогон после выпадения спор начинает увядать, его ножка свободно вынимается из влагалища, при этом поверхность гаустория оказывается не поврежденной - не оторванной, а гладкой (смотреть в лупу).
Вначале спорогон лишен хлорофилла и развивается полностью за счет гаметофита; затем ножка и коробочка становятся ярко-зелеными, спорогон в какой-то мере переходит к самостоятельной ассимиляции; в эпидермисе спорогона появляются настоящие устьица с замыкающими клетками; наконец, в период созревания спорогон теряет зеленую окраску: коробочка становится желтой, а ножка темно-красной.
Для изучения строения коробочки надо приготовить два среза, продольный и поперечный. Сопоставляя их, устанавливаем, что стенка коробочки многослойная и что внутри нее проходит вертикальная колонка, являющаяся продолжением ножки спорогона; верхняя расширенная часть колонки образует тонкую пленочку, называемую эпифрагмой, закрывающую вход в устье коробочки.
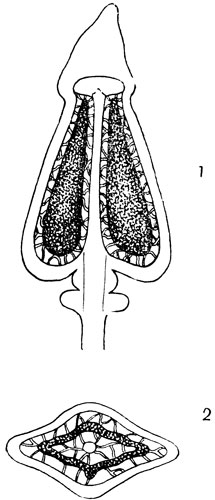
Рис. 54. Коробочка кукушкина льна в прэдольном и поперечном разрезах. В центре разреза - колонка, вокруг - спорангий, в спорангии масса спор
В полости между колонкой и стенкой коробочки помещается спорангий, имеющий форму кольцеобразного складчатого мешка, подвешенного тонкими зелеными нитями к стенке коробочки и к колонке. Спорангий наполнен массой мелких зеленых спор.
Снимем с коробочки ее остроконечную крышечку. Срежем затем бритвой тонкий кольцевой край коробочки вместе с эпифрагмой и рассмотрим его в лупу (лучше 20Х). По краю устья коробочки находится ряд мелких зубцов с закругленными краями, называемый перистомом (околоустьем). Зубцы перистома очень чувствительны к изменению влажности воздуха. В сырую погоду они прижаты к краю эпифрагмы, препятствуя выпадению спор; в сухую погоду, быстро подсыхая, отгибаются, открывая выход для спор. Коробочка, сидящая на длинной и гибкой ножке, раскачивается от ветра, и споры при этом постепенно вытряхиваются из нее. Таким образом, гигроскопичность перистома обеспечивает выпадение спор только в сухую погоду. Мелкие легкие споры подхватываются течениями воздуха и таким образом рассеиваются.
Еще лучше развит перистом на коробочке фунарии влагомерной (Funaria hygrometrica), встречающейся обычно на лесных пожарищах.
На сырой почве споры прорастают в предросток (протонему). В лаборатории производят посев спор мха на влажный песок и содержат под стеклом в прохладном месте (до +15° С) и не на прямом свете.
Микроскопически предростки зеленых мхов очень похожи на нитчатые зеленые водоросли, отличаясь косыми перегородками между клетками. Некоторые ветви протонемы, лишенные хлорофилла, выполняют роль ризоидов. На предростке возникают почки, из которых вырастают взрослые растения.
Сделаем некоторые выводы и обобщения. У всех мохообразных гаметофит связан с водной средой обитания; важнейший процесс, совершающийся на гаметофите, - оплодотворение требует наличия воды как среды для передвижения сперматозоидов. Спорогон, т. е. их спорофит, наоборот, приспособлен к жизни в воздушной среде; споры, образующиеся в спорогоне, распространяются токами воздуха наилучшим образом в сухую погоду.
Рассмотренные формы мохообразных иллюстрируют различную степень усложнения их строения: от простого слоевища печеночников до листостебельного строения у сфагновых и зеленых мхов. Ход эволюции мохообразных не выяснен окончательно. Некоторые считают своеобразную организацию сфагновых мхов вторичным приспособлением их к специфической экологии торфяных болот. По воззрениям некоторых авторов, слоевище печеночников является видоизменением листостебельного побега. По этим взглядам, эволюция мохообразных представляется в такой последовательности: от класса листостебельных мхов к классу печеночников, а в пределах первых - от порядка зеленых мхов к порядку сфагновых.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'