Порядок Эуроциевые (Eurotiales) (И. И. Сидорова)
Этот порядок называют также плектоасковыми (Plectascales) или аспергилловыми (Aspergillales). Он объединяет несколько сотен видов. Плодовые тела эуроциевых - клейстотеции с беспорядочно расположенными внутри прототуникатными сумками - образуются обычно на мицелии на поверхности субстрата или бывают погружены в него. Лишь у немногих представителей этой группы они развиваются в стромах, обычно напоминающих склероции.
Клейстотеции у большинства эуроциевых микроскопические (не более 1-2 мм в диаметре, чаще 100-500 мкм). Исключение составляют только грибы семейства элафомицетовых (Elaphomycetaceae), подземные клейстотеции которых достигают нескольких сантиметров в диаметре.
У некоторых наиболее примитивных эуроциевых клейстотеции отсутствуют и сумки образуются группами на мицелии (например, Byssochlamys, рис. 71); у других перидий клейстотециев очень рыхлый, просвечивающий (например, Amauroascus).
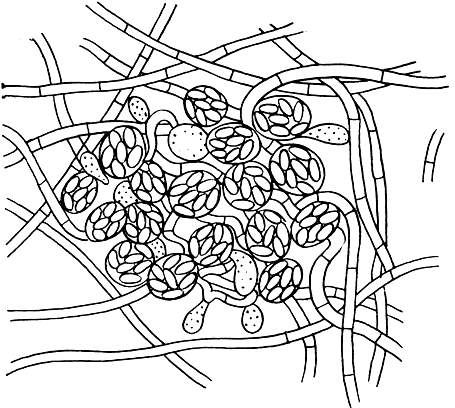
Рис. 71. Биссохламис белоснежный (Byssochlamys nivea)
Перидий клейстотециев имеет разнообразное строение, от рыхлого, паутинистого переплетения гиф, мало отличающихся от вегетативных (например, у Amauroascus, Arachniotus), до плотного псевдопаренхиматического (Elaphomyces granulatus).
Сумки у эуроциевых развиваются на аскогенных гифах различными способами - цепочками по ходу гифы (например, у Talaromyces flavus), из боковых выростов аскогенных гиф (Eupenicillium), по способу крючка (Eurotium, Sartorya). Они прототуникатные, с быстро разрушающейся оболочкой, шаровидные или грушевидные, с 2-8 аскоспорами. Аскоспоры всегда одноклеточные, бесцветные или окрашенные (красные, фиолетовые, коричневые), шаровидные, эллипсоидальные или линзовидные, часто с разнообразной орнаментацией. Освобождение аскоспор происходит пассивно после разрушения оболочки сумки и перидия клейстотеция.
В распространении большинства эуроциевых существенную роль играет бесполое размножение. Лишь немногие представители этого порядка имеют в цикле развития только сумчатую стадию. Наиболее распространены конидиальные спороношения двух типов - алевриоспоры и фиалоспоры (способы их образования см. в разделе о дейтеромицетах). Первые из них толстостенные, одноклеточные (например, у Emmonsiella capsulata) или многоклеточные, часто очень крупные, 1 до 150 мкм длиной (например, у дерматофитов, рис. 75, 76). Если алевриоспоры встречаются и у некоторых гемиаскомицетов, то второй тип конидий - фиалоспоры - впервые появляется именно у эуроциевых. Этот тип конидий характерен и для некоторых групп пиреномицетов, например гипокрейных. Спорогенные клетки - фиалиды - образуются одиночно на гифах мицелия, например у грибов рода эмерицеллопсис (Emericellopsis) или на специализированных конидиеносцах, часто сложного строения: конидиальные стадии пеницилл (Penicillium) и аспергилл (Aspergillus), характерные для некоторых родов семейств эуроциевых (рис. 231).
У эуроциевых наблюдается также аггрегация конидиеносцев - образование коремий, например у пенициллиопсиса (Penicilliopsis), и даже пикнид у пикнидиофоры (Pycnidiophora). Некоторые эуроциевые образуют мелкие шаровидные или продолговатые склероции.
Большинство эуроциевых - сапрофиты на различных субстратах растительного и животного происхождения, в том числе широко распространенные почвенные грибы, например роды эмерицелла (Emericella), сартория (Sartorya), таларомицес (Talaromyces) и многие другие. Некоторые представители этой группы, развиваясь на пищевых продуктах или различных промышленных материалах и изделиях, вызывают их плесневение и порчу (Byssochlamys fulva на фруктовых соках, многие виды пенициллов и аспергиллов). Кератинофильные эуроциевые образуют кератинолитические ферменты и поэтому могут развиваться на субстратах, содержащих кератин (нерастворимый фибриллярный белок),- на перьях, волосах, копытах, рогах (некоторые Gymnoascaceae), принимая участие в их разложении.
Среди эуроциевых есть также грибы, патогенные для человека и животных и вызывающие нередко тяжелые заболевания. Таковы дерматофиты, возбудители глубоких микозов (например, Emmonsiella capsulata).
Родственные эуроциевым несовершенные грибы из формального рода аспергиллус выделяют афлатоксин, вызывающий токсикоз у животных (см. о дейтеромицетах). Большинство используемых в производстве грибов - продуцентов антибиотиков, ферментов, органических кислот также относятся к этому порядку (виды эмерицеллопсис - продуценты антибиотика цефалоспорина С) или близки к эуроциевым (пенициллы и аспергиллы, см. о дейтеромицетах).
Семейство Гимноасковые (Gymnoascaceae)
Это семейство объединяет группу грибов с мелкими примитивными клейстотециями, шаровидными или неправильной формы, образующимися на поверхности субстрата. Их перидий состоит из рыхлого сплетения гиф, напоминающих вегетативные гифы, например у родов арахниотус (Arachniotus) и амауроаскус (Amauroascus), или из плотно переплетающихся и анастомозирующих гиф, часто с утолщенными стенками. На перидии часто образуются придатки различной длины и формы - простые, не отличающиеся от гиф, спирально закрученные у миксотрихум (Myxotrichum), артродерма (Arthroderma), гребневидные у ктеномицес (Ctenomyces) и ветвящиеся. Их форма - важный систематический признак, используемый при разграничении родов этого семейства.
У родов биссохламис (Byssochlamys) и псевдоарахниотус (Pseudoarachniotus) плодовых тел нет и сумки образуются группами или гроздьями на мицелии, напоминая голые сумки гемиаскомицетов. Однако, в отличие от последних, сумки гимноасковых всегда развиваются на аскогенных гифах.
Конидиальная стадия известна не у всех гимноасковых, хотя у многих представителей этого семейства она преобладает в цикле развития. Конидии обычно образуются по типу алевриоспор. Фиалоспоры в этом семействе встречаются редко. Иногда образуются артроспоры, бластоспоры и хламидоспоры.
Гимноасковые грибы - экологически разнообразная группа. Они обитают сапрофитно в почве, на растительных субстратах, на экскрементах животных. Среди гимноасковых широко распространена кератинофилия (Ctenomyces serratus, дерматофиты). Ряд представителей этой группы вызывает заболевания у человека и животных (дерматомикозы, гистоплазмоз).
Наиболее примитивные гимноасковые - грибы из родов биссохламис и псевдоарахниотус - еще не имеют плодового тела. Их шаровидные или овальные сумки образуют неправильную гроздь или пучок на мицелии (рис. 71). Эти роды различаются по характеру конидиального спороношения: у первого образуются фиалоспоры на конидиеносцах типа пециломицес (Paecilomyces); у второго - алевриоспоры или конидиальное спороношение отсутствует.
В роде биссохламис два вида. Буро-желтый биссохламис (Byssochlamys fulva) распространен во всем мире на большом числе разнообразных субстратов - на пищевых продуктах, особенно консервированных фруктах и соках, на различных изделиях и материалах, в почве. На естественных субстратах и в культуре этот гриб образует колонии от бледно-желтого до табачно-коричневого цвета, сначала с обильным конидиальным спороношением - фиалидами с цепочками конидий, расположенными на мицелии или конидиеносцах одиночно или мутовками. Позднее на мицелии образуются аскогоны - короткие завитки гифы, а из них вырастают аскогенные гифы, на которых развиваются грозди сумок с восемью бесцветными аскоспорами (см. рис. 71). Кроме того, в культуре образуются толстостенные хламидоспоры.
Буро-желтый биссохламис имеет экономическое значение как возбудитель биоповреждений различных видов органических материалов. Когда-то этот вид представлял серьезную проблему для консервной промышленности. Он широко распространен в садовых почвах и на гниющих фруктах. Вместе с плодами он попадает на консервные заводы и заражает готовые продукты, вызывая затем их порчу. Гриб очень устойчив ко многим внешним воздействиям, убивающим другие грибы и бактерии. Его аскоспоры сохраняют жизнеспособность при нагревании их в течение 30 мин до 84-87° С, а при консервировании часть их выживает даже при температуре 98° С. Он может развиваться в герметически закрытых консервных банках, так как не требует высокого содержания кислорода. В конидиальной стадии (Paecilomyces varioti) этот вид обнаружен также на бумаге, хлопковой пряже, выделанной коже, разрушение которых он может вызывать.
Второй вид этого рода - белоснежный биссохламис (В. nivea) - распространен в почве, а также встречается на влажных ботанических препаратах, хранящихся в спирте. Грибы этого вида образуют снежно-белые колонии, с возрастом слегка желтеющие. На мицелии развиваются слабо ветвящиеся конидиеносцы с фиалидами, а затем группы шаровидных сумок, иногда, в отличие от желто-бурого биссохламиса, окруженных рыхло расположенными белыми вегетативными гифами (см. рис. 71).
Грибы рода биссохламис хорошо развиваются при повышенных температурах (30-37° С), образуя много сумок. При более низкой температуре (20-24° С) рост более слабый, образуется только конидиальное спороношение.
У грибов рода псевдоарахниотус (Pseudoarachniotus), встречающихся в почве и на экскрементах животных, конидиальное спороношение обычно отсутствует. Аскоспоры часто окрашенные - красные или оранжевые, лимонно-желтые.
Все остальные гимноасковые имеют клейстотеции с перидием. У видов из родов арахниотус (Arachniotus) и амауроаскус (Amauroascus) гифы перидия тонкостенные и напоминают гифы вегетативного мицелия. Перидий слабо развит и представляет очень рыхлое, паутинистое переплетение гиф. Эти роды различаются по окраске аскоспор: у первого они бесцветные или светлоокрашенные, у второго - коричневые или коричнево-фиолетовые. Кроме того, у видов арахниотус стенки сумок очень быстро разрушаются и аскоспоры выходят из клейстотеция через отверстия между гифами перидия.
Наиболее распространенные виды этих родов белоснежный арахниотус (Arachniotus candidus) с белыми шаровидными клейстотециями, часто встречающийся в Европе на экскрементах животных, на птичьих перьях, нередко его можно обнаружить в птичьих гнездах; красный арахниотус (A. ruber) с оранжевыми или красными клейстотециями около 0,5 мм в диаметре, копрофил; бородавчатый амауроаскус (Amauroascus verrucosus) с белыми, после созревания аскоспор темнеющими клейстотециями, обитающий на гниющей коже.
Для родов гимноаскус (Gymnoascus), миксотрихум (Myxotrichum), артродерма (Arthroderma), наницция (Nanizzia) и других характерен перидий из толстостенных гиф, хорошо отличающихся от гиф вегетативного мицелия, а также образование гифами перидия придатков различного строения.
На конском навозе, а также в почве очень часто встречается гимноаскус Рисса (Gymnoascus reessii), образующий белые паутинистые дерновинки, а на них - многочисленные шаровидные желтые, желто-бурые, а иногда оранжевые клейстотеции, часто сливающиеся в корочки. Перидий клейстотециев состоит из толстостенных, обильно ветвящихся под прямым углом, желтоватых или бурых гиф, с прямыми или изогнутыми в виде багра короткими придатками (рис. 72). Конидиальное спороношение отсутствует. Гимноаскус Рисса нередко развивается также на тканях и других материалах и вызывает их порчу.
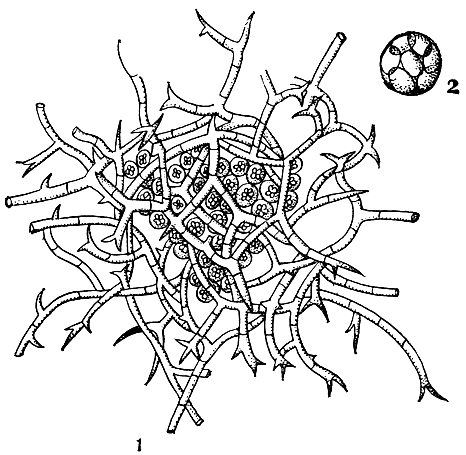
Рис. 72. Гимноаскус Рисса (Gymnoascus reessii): 1 - клейстотеций; 2 - сумка
Род миксотрихум (Myxotrichum) отличается от предыдущего придатками двух типов - короткими шиловидными и длинными, часто спирально закрученными на концах (рис. 73), а также более темным перидием. Виды этого рода встречаются в почве, на экскрементах, различных растительных субстратах, а некоторые - на бумаге и других целлюлозосодержащих материалах (Myxotrichum chartarum).
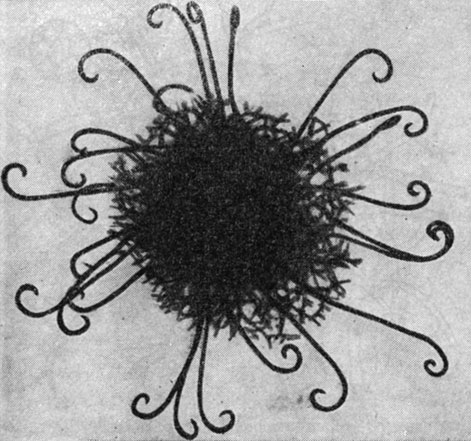
Рис. 73. Миксотрихум (Myxotrichum)
Очень характерные придатки образуются на мелких оранжево-красных клейстотециях пильчатого ктеномицеса (Ctenomyces serratus), распространенного в Европе, Северной и Центральной Америке и Африке сапрофита на птичьих перьях. Эти придатки отходят от толстостенных гиф перидия, слегка изогнуты и состоят из 5-11 толстостенных клеток, каждая из которых образует длинный вырост. Все выросты придатка направлены в одну сторону, и он напоминает гребень (рис. 74).
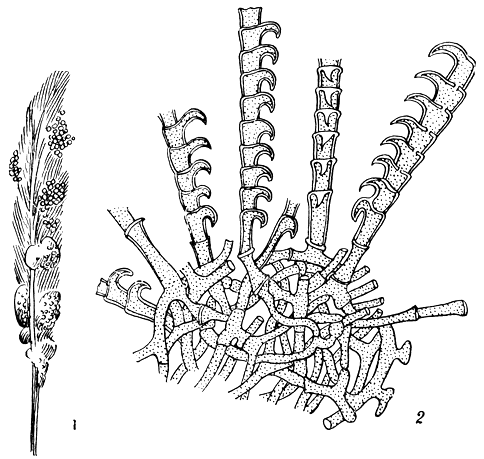
Рис. 74. Ктеномицес пильчатый (Ctenomyces serratus): 1 - клейстотеции на птичьих перьях; 2 - пильчатые придатки клейстотеции
К гимноасковым относится большая группа грибов-дерматофитов, обитающих на волосах, ногтях, коже, а иногда и в тканях и вызывающих дерматомикозы - заболевания человека и многих животных (трихофитию, микроспорию, фавус и т. п.). Дерматофиты, обладая кератинолитическими ферментами и относительной устойчивостью к кожным выделениям животных, занимают своеобразную экологическую нишу, недоступную для большинства других микроорганизмов и относительно свободную поэтому от антагонистов. Кроме того, некоторые дерматофиты образуют антибиотики, например пенициллин у грибов трихофитон (Trichophyton). Антибиотики подавляют развитие сопутствующей микрофлоры.
Дерматофиты - одни из первых обнаруженных патогенных грибов. Начиная с середины прошлого века описано большое число их видов. Однако заболевания, вызываемые ими, были известны задолго до открытия их возбудителей. Например, фавус (парша) был известен уже в течение многих столетий. Длительное время дерматофиты относили к несовершенным грибам, так как у них были известны только бесполые стадии.
В естественных условиях дерматофиты образуют мицелий, распадающийся на артроспоры, а в культуре на питательных средах - обычно обильный и разнообразный рост (различные видоизменения мицелия, макроконидии и микроконидии типа алевриоспор). По характеру макроконидий конидиальные спороношения дерматофитов относят к трем родам: микроспорум (Microsporum) с толстостенными шероховатыми веретеновидными макроконидиями с перегородками, трихофитон (Trichophyton) с тонкостенными гладкими макроконидиями и эпидермофитон (Epidermophyton) с толстостенными гладкими макроконидиями (рис. 75-76). У двух первых родов образуются также микроконидии.
Предположение о связи дерматофитов с гимноасковыми было высказано еще в конце XIX в.
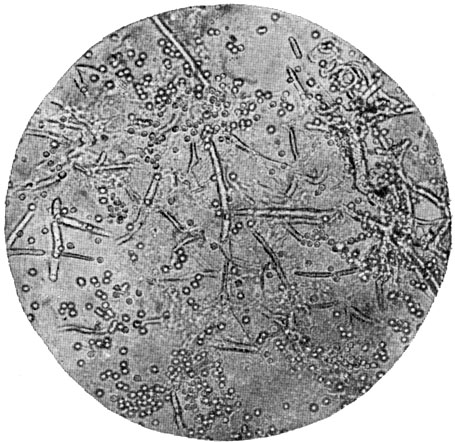
Рис. 75. Гипсовидный трихофитон (Trichophyton gypseum). Конидии

Рис. 76. Микроспорум шерстистый (Microsporum lanosum). Конидии
Эта гипотеза была подтверждена затем А. Наницци, обнаружившим в 1927 г. на кератиновом субстрате в почве клейстотеции гипсовидного микроспорума (Microsporum gypseum). Наконец, в 50-х годах, когда получил распространение метод приманок (волосы, перья), предложенный... Р. Ванбрейзегемом для изучения дерматофитов в почве, были получены сумчатые стадии нескольких видов микроспорумов и трихофитонов, относящиеся к родам наницция (Nanizzia) и артродерма (Arthroderma). Таким образом, были окончательно доказаны родственные связи дерматофитов с гимноасковыми.
Клейстотеции родов артродерма и наницция имеют много общих черт (рис. 77-78). Они шаровидные, 300-700 мкм в диаметре, белые, затем желтеющие. Их перидий состоит из обильно ветвящихся толстостенных бородавчатых или шиповатых гиф, клетки которых часто имеют перетяжки. Придатки - гладкие тонкостенные гифы (прямые, заостренные или в виде спирали с 3-50 витками). Клейстотеции содержат восьмиспоровые сумки с быстро разрушающейся оболочкой, образующиеся по способу крючка. Среди этих грибов встречаются гомоталличные и гетероталличные виды.
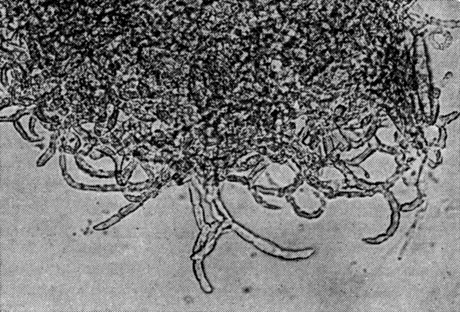
Рис. 77. Наницция загнутая (Nanizzia incurvata) Часть клейстотеция

Рис. 78. Артродерма (Arthroderma). Клейстотеций
Для наницции характерно конидиальное спороношение типа микроспорум, а также заостренные придатки клейстотеция и клетки перидия с несколькими перетяжками. К этому роду относится сумчатая стадия гипсовидного микроспорума - загнутая наницция (Nanizzia incurvata). Конидиальные стадии рода артродерма - это трихофитон и некоторые другие. В перидии имеются многочисленные гантелевидные клетки. Кроме дерматофитов, к артродермам относятся также сапрофиты на растительных остатках, экскрементах, а также сапрофитные кератинофилы.
По степени специализации и приспособления к паразитизму различают несколько групп дерматофитов. Антропофильные дерматофиты вызывают заболевания только у человека. Инфекция передается только от человека к человеку. При попадании в почву грибы этой группы быстро гибнут. Зоофильные дерматофиты (Trichophyton gypseum, Microsporum lanosum и др.) могут вызывать заболевания как у человека, так и у животных, причем инфекция часто передается от домашних животных к человеку (например, М. lanosum, вызывающий микроспорию у кошек и собак). Эта группа дерматофитов часто дает вспышки эпидемического характера, а в некоторых местностях они вызывают до 80-90% всех дерматомикозов (А. Я. Малкина).
Гипсовидный трихофитон (Trichophyton gypseum рис. 79) поражает крупный рогатый скот, овец и других животных, от которых инфекция может передаваться и человеку. Зоофильные дерматофиты паразитируют не только на домашних, но и на диких животных, которые часто не болеют сами, а переносят инфекцию механически. Например, гипсовидный трихофитон был обнаружен у 2,25% обследованных мышей, полевок и землероек в Подмосковье (А. Я. Малкина), а в Великобритании до 20-50% обследованных полевок различных видов оказались носителями персикоцветного трихофитона (Trichophyton persicolor). Гипсовидный микроспорум часто выделяется из почвы и относится к группе геофильных дерматофитов. Наконец, существует группа почвенных кератинофильных грибов (М. cookei, М. ajelloi, Trichophyton terrestre и др.), близких к зоофильным дерматофитам. При искусственном заражении животных некоторые из них слабо патогенны.
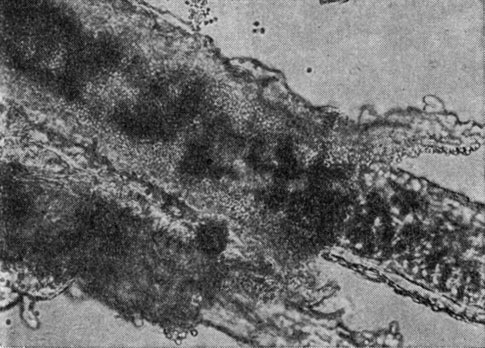
Рис. 79. Волос, пораженный гипсовидным трихофитоном (Trichophyton gypseum)
Сумчатые стадии образуются у почвенных кератинофилов, геофильных и некоторых зоофильных дерматофитов. Антропофильные дерматофиты полностью их утратили.
Большинство дерматофитов - космополиты. Например, гипсовидный микроспорум распространен на всех континентах. Некоторые виды (Microsporum ferrugineum, Trichophyton coucentricum) распространены преимущественно в более теплых районах земного шара, а в умеренной зоне дают лишь эндемические вспышки при ввозе.
Кроме дерматомикозов, гимноасковые могут вызывать и глубокие микозы. В 1972 г. был описан монотипный род эммонсиелла (Emmonsiella) с видом капсульная эммонсиелла (Е. сарsulata) - сумчатой стадией капсулъной гистоплазмы (Histoplasma capsulatum), вызывающей у человека гистоплазмоз - тяжелое поражение ретикуло-эндотелиальной системы, часто со смертельным исходом. Это заболевание было обнаружено в начале нашего века в зоне Панамского канала, но его возбудитель был неправильно отнесен сначала к простейшим. Гистоплазмоз распространен преимущественно в странах с мягким климатом. Его локальные очаги известны в некоторых штатах США, странах Южной Америки, Азии, Африки и Европы.
Источником инфекции многие исследователи считают почву. Капсульная гистоплазма часто выделяется из почвы и воды эндемических районов, однако значительно чаще она встречается в экскрементах различных животных - скворцов, летучих мышей, кур и др. Известны случаи, когда гистоплазмозом заболевали группы спелеологов после посещения пещер, содержащих большое количество гуано летучих мышей.
Развитие капсульной гистоплазмы, как и многих других возбудителей глубоких микозов, происходит в два этапа. В организме человека она развивается в паразитической - дрожжеподобной фазе, образуя ее также на средах специального состава (например, с кровью) при +37 °С. На средах иного состава и при более низкой температуре развивается сапрофитная фаза гриба - мицелий с толстостенными одноклеточными алевриоспорами (рис. 80). Заражение вызывают конидии сапрофитной фазы.
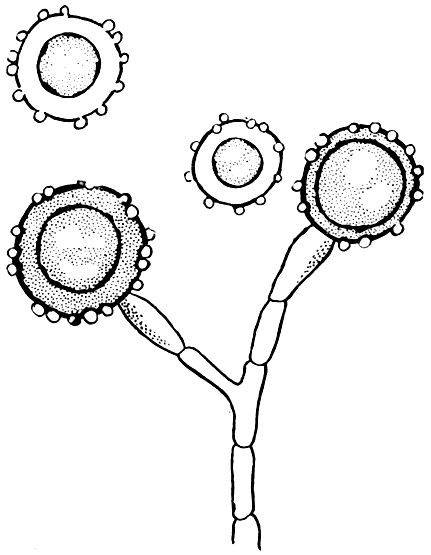
Рис. 80. Гистоплазма капсульная (Histoplasma capsulatum). Конидии
Клейстотеции гриба белые, позднее буроватые, шаровидные, неправильно звездчатые, 80-250 мкм в диаметре. Перидий состоит из гиф двух типов - спирально скрученных и отходящих от них ветвящихся волнистых гиф.
Семейство Эуроциевые (Еurotiасеае)
К семейству эуроциевых относятся плектомицеты с хорошо развитыми клейстотециями, перидий которых псевдопаренхиматический или имеет четко выраженную гифенную структуру.
За немногими исключениями, грибы из этого семейства - широко распространенные сапрофиты. Они обитают в почвах различных климатических областей, а также на разнообразных субстратах растительного, реже животного происхождения, на которых они образуют плесени. Некоторые эуроциевые - термофилы и развиваются на разнообразных саморазогревающихся субстратах (например, в компосте, влажном сене и др.) при температуре 30-60 °С. Отдельные виды вызывают заболевания теплокровных животных и растений.
Человек в своей деятельности часто встречается с грибами из этой группы. Это не только многочисленные и всем хорошо известные зеленые, голубые и черные плесени на разнообразных пищевых продуктах. Они развиваются также на различных промышленных изделиях и материалах. В благоприятных для развития условиях, особенно в тропиках, они могут за короткий срок вызывать разрушение тканей, кожи, различных синтетических материалов (например, электроизоляции), ускорять процессы коррозии металлов, повреждать приборы, оптику и многие другие изделия. С другой стороны, некоторые из эуроциевых и близких к ним несовершенных грибов из формальных родов аспергилл (Aspergillus) и пеницилл (Penicillium) широко используют в микробиологической промышленности как продуценты антибиотиков (пенициллины, цефалоспорин С и др.), ферментов и органических кислот, а также для приготовления некоторых пищевых продуктов - сыров (рокфор, камамбер) и соусов.
В распространении большинства эуроциевых большую роль играет конидиальная стадия. Нередко она преобладает в цикле развития, а клейстотеции образуются лишь спорадически. У большой группы грибов, связанных по происхождению с эуроциевыми, сумчатые стадии утрачены полностью и конидиальное спороношение - единственный способ размножения. К этой группе относятся такие широко распространенные почвенные грибы, как многие пенициллы и аспергиллы, а также некоторые виды из рода акремониум (Acremonium). Они принадлежат к классу дейтеромицетов, или несовершенных грибов.
Преобладающий у эуроциевых тип конидий - фиалоспоры. Они образуются на фиалидах в базипетальных цепочках, а иногда собираются в ложные головки. Фиалиды расположены поодиночке на недифференцированных гифах мицелия, например у рода эмерицеллопсис (Emericellopsis) в конидиальной стадии типа акремониум, но значительно чаще - на хорошо развитых конидиеносцах сложного строения (конидиальные стадии типа пеницилл и аспергилл, характерные для многих родов эуроциевых). Конидиеносцы обычно одиночные, однако у некоторых грибов из этого семейства они объединяются в коремии, например у пенициллиопсис (Penicilliopsis). В 1955 г. был описан род эуроциевых с конидиями, развивающимися в пикнидах,- пикнидиофора (Pycnidiophora).
Образованию клейстотециев предшествует дикарионтизация, которая может происходить у эуроциевых несколькими путями. У одних представителей этого семейства наблюдается типичный для высших аскомицетов половой процесс (например, у Monascus purpureus). Однако у многих эуроциевых происходит его морфологическая редукция. У ползучего эуроциума (Eurotium repens) и желтого таларомицеса (Talaromyces flavus) антеридии образуются, но не функционируют, а у неосартории Фишера (Neosartorya fischeri) они вообще отсутствуют. В этих случаях в дикарионы объединяются ядра самого аскогона. Наконец, у лежачей эмерицеллы (Emericella nidulans) и ряда других видов гаметангии не образуются и дикарионтизация происходит соматогамно, в результате слияния двух обычных клеток вегетативного мицелия.
Клейстотеции эуроциевых обычно достигают 100-500 мкм в диаметре и заметны невооруженным глазом в виде маленьких шариков. Часто они ярко окрашены (желтые, оранжевые), но могут также быть белыми, светлоокрашенными или темными. У большинства видов этого семейства они образуются на мицелии на поверхности субстрата, но у некоторых его представителей имеются небольшие стромы, часто по твердости напоминающие склероции, и клейстотеции развиваются внутри них, например у рода петромицес (Petromyces). Перидий клестотециев образуется обычно из переплетающихся гиф, но у некоторых видов (Pseudoeurotium multisporum и др.) он имеет тканевое строение, развиваясь в результате многократного деления одной или нескольких клеток гифы.
Аскоспоры эуроциевых всегда одноклеточные, бесцветные или разнообразно окрашенные (у эмерицеллы - фиолетовые, красные, пурпурные, у эмерицеллопсиса - коричневые), овальные или линзовидные, часто с разнообразной орнаментацией - шероховатые, с экваториальной бороздкой, ребрами, крыловидными выростами.
Большую группу эуроциевых составляют грибы, конидиальные стадии которых относятся к формальным родам несовершенных грибов: пепицилл (Penicillium) и аспергилл (Aspergillus). Эти два рода объединяют многочисленные виды грибов, широко распространенных в почвах всего земного шара от Арктики до тропиков, а также на различных субстратах растительного происхождения. У многих из них известны только конидиальные стадии, такие виды отнесены к несовершенным грибам и описаны в соответствующем разделе.
Однако у некоторых пенициллов и аспергиллов, по традиции также относимых к несовершенным грибам, известны сумчатые стадии, принадлежащие к различным родам эуроциевых.
Согласно Международному кодексу ботанической номенклатуры, основным наименованием плеоморфных грибов является наименование их совершенной стадии. Поэтому мы рассматриваем эти грибы в настоящем разделе.
Для формального рода аспергилл характерны простые конидиеносцы, на вершине вздутые в виде пузыря различной формы. На нем располагаются фиалиды, образующие цепочки одноклеточных конидий. У некоторых аспергиллов фиалиды располагаются не на самом пузыре, а на образующихся на нем профиалидах (рис. 231). Подробное описание конидиального аппарата у этого рода приведено в разделе о дейтеромицетах. Конидиальные стадии этого типа известны у девяти родов эуроциевых. Интересно отметить, что виды одного рода обычно имеют конидиальные стадии, относящиеся к одной или близким группам рода аспергилл.
Наиболее обширный род этой группы - эуроциум (Eurotium) - включает 18 видов. Его представители часто встречаются в природе на разнообразных медленно разлагающихся растительных субстратах. Они образуют зеленые, желтоватые или красно-желтые плесени, в зависимости от условий среды (температуры, влажности) и, соответственно, преимущественного развития конидиальной или сумчатой стадии (табл. 16). Клейстотеции грибов этого рода обычно шаровидные, очень мелкие (50-175 мкм в диаметре), желтые, с однослойным перидием, покрыты рыхлой сетью гиф с желтоватыми или красноватыми гранулами. Оболочки сумок разрушаются очень рано, и зрелые клейстотеции содержат массу бесцветных или желтоватых линзовидных аскоспор с экваториальной бороздкой. Конидиальные стадии относятся к группе Aspergillus glaucus.
Первый крупный вид этого рода - гербарный эуроциум (Eurotium herbariorum) - был обнаружен Г. Ф. Липком в 1809 г. на сухом гербарном образце. Его связь с конидиальной стадией из рода Aspergillus - сизым аспергиллом (A. glaucus) - была доказана значительно позднее, в 1854 г., А. де Бари.

Таблица 16. Колонии эуроциевых грибов: вверху слева - эуроциум ползучий (Eurotium repens); вверху в середине - неосартория Фишера (Neosartorya fischeri); вверху справа - петромицес чесночный (Petromyces alliaceus); в середине справа - таларомицес желтый (Talaromyces flavus); внизу слева - эмерицеллопсис почвенный (Emericellopsis terricola); внизу справа - конидиальная стадия гриба из рода эмерицеллопсис (Emericellopsis)
В почве эуроциумы встречаются в небольшом количестве. Обычный субстрат многих видов из этого рода - хранящиеся растительные продукты. Многие из его представителей - ксерофиты, развивающиеся в условиях пониженной влажности, неблагоприятных для роста других грибов. Ползучий эуроциум (Eurotium repens), например, вызывает плесневение зерна и многих других продуктов при влажности 13-15%. В результате его развития освобождается вода и на зерне начинают расти другие плесневые грибы. Ползучий эуроциум очень быстро распространяется в хранилищах, так как обитающие там долгоносики и клещи, охотно питаясь его гифами и спорами, разносят их по всем помещениям.
Большое значение имеют эуроциумы и как возбудители биоповреждений различных промышленных изделий и материалов. Развиваясь при минимальных влажности и питании, они вызывают разложение текстиля, целлофана, резины, пластмасс (Е. repens, Е. amstelodami). Грибы этого рода (например, Е. tonophilum) встречаются даже на стеклах оптических приборов, вызывая их порчу. Они ускоряют также процессы коррозии металлов, вероятно, вследствие образования большого количества органических кислот.
Эуроциумы обычно осмофильны и могут развиваться на средах с высоким осмотическим давлением, например с повышенным содержанием сахара (20% и более). Ползучий эуроциум часто встречается на заплесневелых джемах и вареньях, где образует обильные конидии и клейстотеции. Галофилъный эуроциум (Е. halophilicum) плохо растет на средах с низким осмотическим давлением. Для его нормального развития необходимо высокое содержание в среде сахара (более 20-40%) или эквивалентной молярной концентрации хлористого натрия. Е. tonophilum хорошо растет и образует клейстотеции на средах с еще большим осмотическим давлением, содержащих до 60% сахара.
Второй крупный род эуроциевых с конидиальной стадией Aspergillus - эмерицелла (Emericella) - объединяет 13 видов. Они имеют шаровидные, довольно крупные клейстотеции (300- 400 мкм в диаметре), обычно ярко-желтые, окруженные массой крупных толстостенных клеток. Встречаются в почвах различных районов земного шара, довольно часты на различных растительных материалах.
Один из наиболее распространенных видов - лежачая эмерицелла (Emericella nidulans, конидиальная стадия A. nidulans) образует широко растущие колонии, окраска которых варьирует от зеленой до ярко-желтой, в зависимости от степени развития конидий и клейстотециев. Клейстотеции желтые, хорошо видны невооруженным глазом. Лежачая эмерицелла обычно встречается на различных растительных субстратах, в почвах умеренной зоны и субтропиков, иногда развивается в дыхательных путях теплокровных животных. В генетике грибов этот вид занимает второе место после видов рода нейроспора (Neurospora).
Другие грибы рода эмерицелла встречаются преимущественно в почвах более теплых и сухих районов: эмерицелла мелкоморщинистая (Е. rugulosa), четырехлинейная (Е. quadrilineata) или в тропических лесных почвах - эмерицелла гетероталличная (Е. heterothallica).
Для грибов рода неосартория (Neosartorya) характерны шаровидные, обычно белые клейстотеции, в зрелости окруженные рыхлой сетью стерильных гиф, придающих им вид маленьких ватных шариков (табл. 16). Их перидий состоит из слоя псевдопаренхимы, толщиной в несколько клеток. Аскоспоры бесцветные, с экваториальными гребнями. Конидиальные стадии - из группы Aspergillus fumigatus. Наиболее распространенный вид этого рода - неосартория Фишера (N. fischeri) - обитает в почве и на различных органических субстратах. При температуре ниже +25 °С он образует желтовато-белые колонии с обильными белыми клейстотециями (150-200 мкм в диаметре). Если те же культуры этого гриба выращивать при повышенных температурах (30-37 °С), они образуют преимущественно конидии, придающие колониям голубовато-зеленую окраску. Клейстотеции в этих условиях не образуются или бывают немногочисленны (табл. 16).
В отличие от перечисленных родов для грибов монотипного рода петромицес (Petromyces) характерны клейстотеции, образующиеся не на мицелии, а в твердых стромах, напоминающих склероции. Единственный вид этого рода - чесночный петромицес (P. alliaceus), его конидиальная стадия - A. alliaceus. Гриб обнаружен в почвах Европы, Азии, Америки и Австралии. Он встречается также на поврежденных луковицах чеснока, лука и других растений. В культуре гриб образует быстро растущие колонии с многочисленными склероциеподобными стромами, расположенными концентрическими зонами. Стромы обычно эллиптические, 1-3 мм длиной, из толстостенных клеток. Их окраска варьирует от серебристо-серой до черной (табл. 16). Клейстотеции образуются внутри этих стром по 1-8 в каждой, в зависимости от ее размера. Их созревание происходит очень медленно, в зрелости они заполняют весь склероций. Тонкая собственная оболочка клейстотециев быстро разрушается, и строма выглядит как один клейстотеций с толстой оболочкой (до 200 мкм толщиной). У грибов другого строматического рода - склероклейста (Sclerocleista) - каждая строма содержит лишь один клейстотеций.
Виды формального рода пеницилл, к которому относятся конидиальные стадии трех родов эуроциевых - эупенициллиум (Eupenicillium), таларомицес (Talaromyces) и хамигера (Hamigera),- образуют конидиеносцы, имеющие вид кисточки (рис. 231). Их строение разнообразно: кисточки состоят из одной мутовки фиалид на конидиеносце или они двухъярусные и состоят из метул и расположенных на них фиалид, наконец, конидиеносец может ветвиться, обычно несимметрично. Сумчатые стадии известны у немногих видов этого формального рода.
У видов рода эупенициллиум клейстотеции шаровидные или неправильной формы, развиваются из псевдопаренхиматических, иногда очень твердых и напоминающих склероции стром. Созревание клейстотеция идет от центра стромы, а его бесцветный или окрашенный перидий состоит из толстостенных клеток стромы. Сумки образуются на боковых веточках аскогенных гиф, одиночно или цепочками. Аскоспоры обычно линзовидные, бесцветные, желтоватые или бледно-коричневые, часто с экваториальным гребнем или бороздкой (табл. 15). В этом роде сейчас насчитывается более 30 видов, обитающих в почве, но часто встречающихся на различных растительных субстратах. У одного из наиболее распространенных видов этого рода - эупенициллиума Брефелъда (Е. brefeldianum, конидиальная стадия - P. brefeldianum) - клейстотеции обычно развиваются из параплектенхиматических, иногда слабосклероциальных стром. Они кремовые или песочного цвета, 100-200 мкм в диаметре. Их созревание происходит за 10-14 дней. Этот вид обитает преимущественно в южных почвах (Центральная Америка, Африка), но встречается также в почвах умеренной зоны.
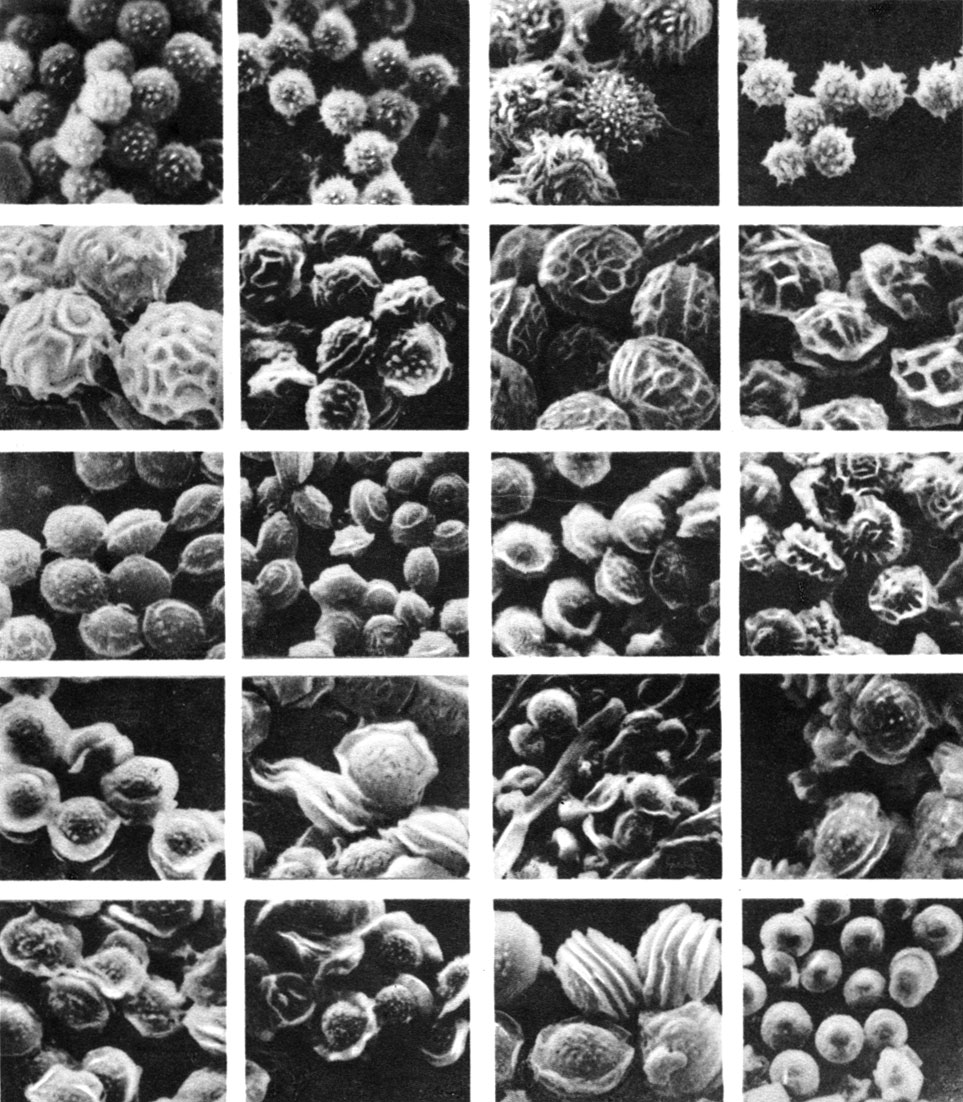
Таблица 15. Аскоспоры эупенициллов (Eupenicilliom) (фото в сканиоуемом микроскопе)
У других видов рода эупенициллиум, например у мелкого эупенициллиума (Е. parvum, конидиальная стадия - P. parvum), развитие клейстотециев длится не менее 20-30 дней, а у корковидного (Е. crustaceum, конидиальная стадия - P. kewense) и эупенициллиума Шира (Е. shearii, конидиальная стадия - P. shearii) склероциевидные клейстотеции созревают за 4-6 недель, а иногда вообще не образуют сумок.
Пеницилл Тома (Penicillium thomii) - гриб, часто встречающийся в лесных почвах, а также на древесине и других растительных субстратах. Он образует розовые склероции, очень похожие по форме и консистенции на молодые клейстотеции эупенициллиумов мелкого, Шира и корковидного. Однако они никогда не содержат сумок и представляют, вероятно, рудиментарные клейстотеции. Это подтверждается и тем, что у эупенициллов иногда клейстотеции недоразвиваются и не образуют сумок.
Род таларомицес объединяет 13 видов с мелкими шаровидными или неправильными клейстотециями с неограниченным ростом. Яркие желтые или оранжево-желтые клейстотеции образуются в изобилии и придают характерную желтую окраску колониям грибов из этого рода (табл. 16). Их перидий развит в разной степени, от очень тонкого, просвечивающего (Т. flavus, Т. wortmannii) до плотного (Т. thermophilus). Сумки образуются цепочками из клеток аскогенных гиф, их оболочки быстро разрушаются.
Виды этого рода обитают в почве и на разнообразных органических материалах. Желтый таларомицес (Т. flavus, конидиальная стадия - P. vermiculatum) и m. Вортмана (Т. wortmannii, конидиальная стадия - P. wortmannii) - космополиты, широко распространенные в почве и часто вызывающие биоповреждения различного оборудования и материалов, особенно в тропиках и субтропиках.
Интересны термофильные виды рода таларомицес - термофильный (Т. thermophilus, конидиальная стадия - P. dupontii) и Эмерсона (Т. emersonii, конидиальная стадия - Р. emersonii). Они развиваются в почве, компосте, саморазогревающемся влажном сене и других субстратах в границах температуры 25-60 °С. При 45 °С они быстро растут и образуют обильное конидиальное спороношение. У первого из них клейстотеции развиваются только в частично анаэробных условиях, на стерильном зерне. Они шаровидные, около 1 мм в диаметре, серые или коричневатые. Клейстотеции таларомицеса Эмерсона типичного для этого рода желтого или оранжевого цвета, более мелкие (50-300 мкм в диаметре), обычно сливаются и образуют слой или корочку на поверхности среды. Их развитие происходит в температурных границах 35-50 °С, в аэробных условиях.
К термофильным грибам относится также оранжевый термоаскус (Thermoascus aurantiacus), образующий быстро растущие кремово-белые колонии, на которых сначала развивается конидиальное спороношение типа пециломицес (Paecilomyces), которое быстро сменяется сумчатым - ярко-оранжевыми или кирпично-красными клейстотециями неправильной формы. Этот вид часто развивается на саморазогревающихся субстратах. Он хорошо переносит анаэробные условия в течение нескольких дней.
Род эмерицеллопсис (Emericellopsis) объединяет почвенные грибы с маленькими шаровидными клейстотециями, одетыми тонким, просвечивающим перидием из двух или более слоев клеток и характерными коричневыми аскоспорами с крыловидными придатками. При росте на питательных средах виды этого рода образуют медленно растущие колонии, на свету приобретающие розовую или оранжевую окраску вследствие образования пигментов из группы каротиноидов. Сначала развивается конидиальное спороношение типа акремониум,- одиночные фиалиды с фиалоспорами, соединенными слизью в ложные головки. Клейстотеции образуются на поверхности среды, на гифах воздушного мицелия, или погружены в среду. Их размеры от 15 до 400 мкм, самые мелкие содержат только по 1-2 сумки. Аскоспоры овальные, коричневые, с крыловидными гребнями различной формы и размера, часто проходящими по всей споре, от полюса до полюса. После массового образования клейстотециев колонии темнеют, так как коричневые аскоспоры просвечивают через тонкий перидий (табл. 16).
Представители рода эмерицеллопсис встречаются в различных типах почв в Европе, Азии, Африке и Северной Америке. Их можно обнаружить как в окультуренных, так и неокультуренных почвах, например лесных. Они часто обитают в сильно увлажненных почвах - в мангровых болотах, торфяниках, в иле (например Е. minima, Е. glabra).
Некоторые виды этого рода (Е. glabra, Е. terricola) - активные продуценты антибиотика цефалоспорина С, близкого по структуре и свойствам к пенициллинам, но менее активного, чем последний. На основе этого антибиотика в последние годы были получены полусинтетические производные, которые не только превосходят по активности исходный препарат, но и действуют на те группы организмов, которые устойчивы к действию пенициллина,- грамотрицательные бактерии и устойчивые к пенициллину стафилококки. Полусинтетические цефалоспорины - цефалотин и цефалоридин - выпускаются промышленностью и используются в медицинской практике.
Виды из рода монаскус (Monascus), вызывающие плесневение различных растительных продуктов, образуют колонии с характерной красной или пурпурной окраской, обусловленной присутствием пигмента монасцина. На концах гиф мицелия у грибов из этого рода образуются шаровидные клейстотеции с перидием из рыхло переплетающихся гиф. Стенки сумок быстро разрушаются, и зрелый клейстотеций содержит массу свободных аскоспор. Наиболее известный вид - красный монаскус (М. ruber) - встречается на гнилых яблоках и других растительных субстратах. Другой вид этого рода - пурпурный монаскус (М. purpureus) - используется в Юго-Восточной Азии для получения окрашенного риса. Гриб выращивают на рисе, из которого затем готовят порошок и используют его для приготовления соусов. В благоприятных для роста условиях этот гриб вызывает плесневение каучука.
Некоторые тропические эуроциевые образуют хорошо дифференцированные стромы, на которых образуются довольно крупные клейстотеции. Так, у рода батистия (Batistia) клейстотеции образуются на псевдопаренхиматических ножках и достигают 3-4 мм в диаметре. У грибов рода пенициллиопсис стромы шаровидные или неправильно лопастные, 2-7 мм в диаметре, одиночные или собранные гроздьями, на коротких ножках. Представители этого рода встречаются на плодах и семенах различных растений в тропической Африке, Юго-Восточной Азии, Бразилии.
Семейство Элафомицетовые (elaphomycetасеае)
Грибы этого семейства образуют крупные подземные клейстотеции с толстым прочным перидием. Шаровидные или грушевидные 8-споровые сумки расположены в них в беспорядке, обычно группами, разделенными стерильными "жилками". После созревания сумок их оболочка разрушается и аскоспоры заполняют клейстотеций в виде порошка.
Элафомицетовые вследствие сходства условий обитания имеют конвергентное сходство с трюфелевыми, поэтому некоторые микологи переносят их в этот порядок. Однако по развитию плодовые тела элафомицетовых отличаются от подземных апотециев трюфелевых, представляя типичные клейстотеции.
Центральный род семейства - элафомицес (Elaphomyces) - объединяет около 25 видов, распространенных в Европе, Азии, Северной Америке и Австралии. Для многих из них установлено образование микоризы с различными деревьями. Клейстотеции элафомицетов образуются в почве на глубине нескольких сантиметров, шаровидные или клубневидные, 1-5 см в диаметре. В зрелости они содержат порошок аскоспор, обычно темного цвета. Перидий их плотный, с гладкой или бугорчатой поверхностью, сверху часто покрыт коркой, состоящей из гиф (рис. 81). Иногда в нее вплетаются концы корней деревьев, окружающих клейстотеции. У некоторых видов корка легко снимается.
Связь элафомицетов с корнями деревьев была обнаружена еще К. Виттадини в 1831 г., однако позднее ее объясняли паразитизмом этих грибов на корнях (Л. Тюлянь, Ш. Тюлянь, Э. Будье). Лишь в 1882 г. Г. Гибелли доказал, что они образуют микоризу.
Распространение видов рода элафомицес изучено слабо, так как их плодовые тела образуются в почве и их трудно обнаружить. При их поиске хороший ориентир представляют стромы грибов рода кордицепс (Cordyceps), паразитирующих на клейстотециях элафомицетов. Они поднимаются над поверхностью почвы, и их легко заметить. Часто можно также найти клейстотеции элафомицетов, раскопанные оленями, кабанами и другими животными. Плодовые тела этих грибов имеют очень острый запах, олени легко их находят и охотно поедают. Эти грибы в ряде стран Европы и Америки даже получили название "олений трюфель". Для человека они несъедобны.
Наиболее распространенный вид - олений трюфель, или зернистый элафомицес (Elaphomyces granulatus),- часто встречается осенью в хвойных лесах, особенно на песчаной почве. Его клейстотеции шаровидные, желто-коричневые, размером 1-4 см, с мелко бородавчатой поверхностью (рис. 81). Мицелий гриба, характерного желтого цвета, оплетает корни деревьев. Он образует микоризу с хвойными (сосна, ель) и некоторыми лиственными деревьями.
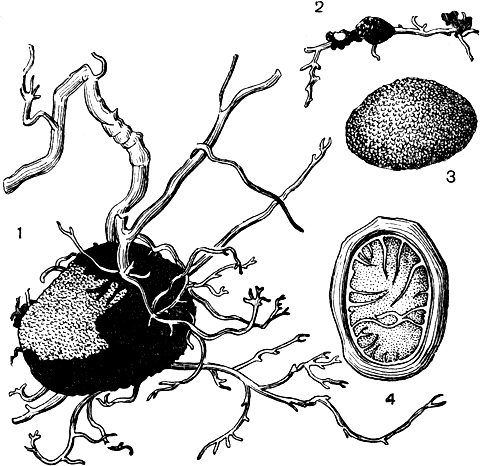
Рис. 81. Элафомицес зернистый (Elaphomyces granulatus): 1, 2 -клейстотеций среди корней дерева; 3 - внешний вид клейстотеция; 4 - разрез клейстотеция
В горных лесах грибы этого вида и сетчатого элафомицеса (Е. reticnlatus) встречаются на высоте до 2700-2800 м над уровнем моря.
Пестрый элафомицес (Е. variegatus) имеет желтовато-серые или черновато-бурые клейстотеции с более бугорчатой поверхностью, чем предыдущий. Он образует микоризу с буком, дубом и хвойными. Реже встречается угольно-черный элафомицес (Е. anthracinus) с черным перидием, образующий микоризу с березой и буком. Он распространен в Центральной Европе и некоторых районах Северной Америки.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'