Порядок Эндомпцетовые (Еndomycetales) (И. И. Сидорова)
Семейство Эндомицетовые (Еndomycetaceae)(И. И. Сидорова)
Большинство представителей эндомицетовых грибов развивается как сапрофиты на поверхности плодов и вегетативных частей растений, в истечениях деревьев, в нектаре цветков, широко распространены в почвах. Некоторые грибы встречаются в ассоциации с насекомыми. Паразитов в этой группе немного (например, Spermophthora gossypii, паразитирующий в коробочках хлопчатника, Endomyces decipiens - паразит на пластинках некоторых агариковых). Большое практическое значение имеют дрожжи - возбудители спиртового брожения, а также продуценты некоторых витаминов (например, Eremothecium ashbyi - продуцент рибофлавина, образующий до 6 г этого витамина на 1 л культуры).
У представителей небольшого по объему семейства диподасковых (Dipodascaceae), включающего 3 рода, на естественных субстратах и в культуре на питательных средах образуется хорошо развитый, ветвящийся мицелий, а на нем - цилиндрические многоспоровые сумки.
Виды этого семейства обитают в слизистых истечениях растений, на древесине, в почве, в ассоциациях с насекомыми, с кольчатыми червями и не имеют практического значения. Но эта группа представляет большой теоретический интерес как возможное связующее звено между аскомицетами и их гипотетическими предками, близкими к современным зигомицетам. Наиболее детально в этом плане изучен род диподаскус (Dipodascus).
Беловатый диподаскус (Dipodascus albidus) был найден в 1892 г. Г. Лагерхеймом в слизистых истечениях растения из семейства бромелиевых. Позднее этот вид был обнаружен в различных частях Европы, Азии и Америки в экскудатах на пнях различных деревьев (березы, граба, бука и др.). Хорошо развитый мицелий этого гриба состоит из крупных (до 100 мкм длиной) многоядерных клеток. На нем сначала развивается бесполое спороношение - многоядерные артроспоры в цепочках, а затем многоядерные гаметангии, различающиеся по размеру. Сливаясь, они образуют зиготу, сразу же развивающуюся в удлиненную, суженную на вершине сумку (рис. 53).
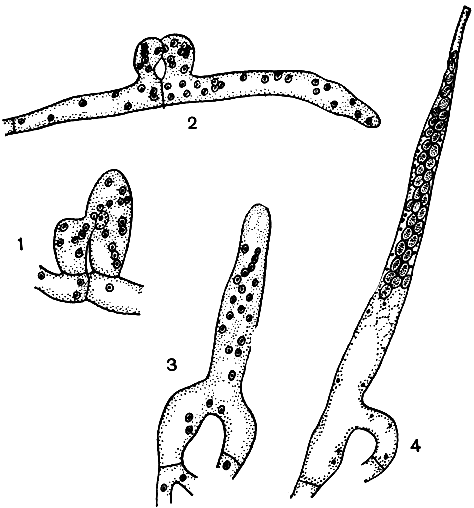
Рис. 53. Диподаскус беловатый (Dipodascus albidus): 1, 2 - слияние гаметангиев; 3 - молодая сумка с гаплоидными ядрами; 4 - зрелая сумка с многочисленными аскоспорами
Освобождение аскоспор происходит пассивно. Они окружены слизистой обверткой, при ее набухании сумка растягивается и разрывается на вершине. Аскоспоры выходят из сумки и собираются на ее вершине в шарик, склеенный слизью. Они прорастают через 24-36 ч в капле воды или на подходящем субстрате.
У двух других видов - одноядерного диподаскуса (Dipodascus uninucleates), обнаруженного в Канаде и США на куколках фруктовой дрозофилы, и собранного диподаскуса (Dipodascus aggregates), встречающегося в Европе и Северной Америке в личиночных галереях короедов, - мицелий состоит из одноядерных клеток. Гаметангии также одноядерны.
Таким образом, развитие спор полового размножения у диподасковых имеет большое сходство с наблюдаемым у зигомицетов. Как и у зигомицетов, здесь может происходить слияние многоядерных гаметангиев (Dipodascus albidus), сменяющихся в процессе эволюции одноядерными (Dipodascus uninucleates). Однако при слиянии многоядерных гаметангиев диподасковых не происходит множественной кариогамии (слияние ядер), а сливаются только два ядра. Здесь следует отметить, что уже у некоторых зигомицетов (род Endogone) при гаметангиогамии сливаются только два ядра, а остальные ядра многоядерных гаметангиев дегенерируют. Сумка аскомицетов гомологична зародышевому спорангию зигомицетов, у диподасковых же их сходство увеличивается еще и тем, что количество аскоспор в сумке может достигать нескольких десятков и не фиксировано строго, как у остальных аскомицетов.
Однако у диподасковых есть и существенные отличия от зигомицетов. Зигота перед прорастанием не переходит в состояние покоя, а сразу превращается в сумку. Образование аскоспор происходит по способу, типичному для аскомицетов ("свободное образование клеток").
Переходный характер семейства диподасковых в известной мере подтверждается результатами биохимических исследований, в частности изучением первичных структур ДНК.
Собранный диподаскус образует интересную в биологическом отношении ассоциацию с некоторыми короедами (Л. Р. Батра, 1959). Запах гриба привлекает жуков, а клейкая обвертка аскоспор обеспечивает их перенос жуками. Этот вид обнаружен как в личиночных ходах короедов, так и в почве (под пораженными короедами деревьями), куда его споры вымываются дождем. Предполагают, что зимующей стадией гриба являются аскоспоры, которые сохраняют жизнеспособность в почве более 15 месяцев.
В семействе эндомицетовых (Endomycetaceae), в отличие от предыдущего, число спор в сумке строго фиксировано и равно восьми или меньше. Среди этой группы преобладают сапрофиты на богатых сахаром субстратах, отдельные виды паразитируют на грибах и животных. Некоторые эндомицетовые грибы используют в странах Азии и Африки для брожения, но в целом группа не имеет практического значения.
Мицелий у представителей этого семейства может состоять из хорошо развитых гиф (род Eremascus), частично распадаться на артроспоры (род Endomyces) либо почковаться (род Endomycopsis).
У грибов рода эремаскус (Eremascus) гаметангиогамия сходна с таковой у диподасковых. Гаметангии представляют отростки клеток мицелия, содержащие по одному ядру, они сливаются, зигота отделяется перегородками от оснований гаметангиев и превращается в восьмиспоровую сумку. У эндомицеса Магнуса (Endomyces magnusii), обитающего в соке деревьев, образование сумок в типе такое же, но гаметангиогамия гетерогамная. Некоторые эндомицетовые образуют сумки апомиктически, например Endomyces decipiens, паразитирующий на пластинках осеннего опенка, или весенний эндомикопсис (Endomycopsis vernalis), часто встречающийся в соке березы, а также в почве и образующий в культуре большое число дрожжеподобных клеток.
Endomyces geotrichum, встречающийся часто в почве и на различных органических субстратах, образует обильные артроспоры (бесполое спороношение - Geotrichum candidum), гомоталличен, но самостерилен. Его сумки содержат по одной аскоспоре. Этот гриб может поражать плоды различных растений, например вызывать гниль плодов цитрусовых и томатов. Его мицелий интенсивно развивается в ткани плодов и через 5 дней образует обильные артроспоры, а иногда и сумки с аскоспорами.
У распространенного в Китае эндомицеса Линднера (Endomyces lindneri), сбраживающего сахарозу и многие моносахариды, зигота не всегда образует сумку сразу, а иногда дает короткую, даже ветвящуюся гифу, на которой образуются сумки. Гифа напоминает по функции аскогенные гифы высших аскомицетов, но, в отличие от них, содержит диплоидные ядра, а не дикарионы.
Увеличение числа сумок, образующихся из одной зиготы, может происходить у эндомицетовых и другим путем. У одонтиевого мириогониума (Myriogonium odontiae), паразитирующего на плодовых телах базидиомицета одонтия (Odontia), впервые у аскомицетов встречается образование сумки по способу крючка.
Таким образом, семейство эндомицетовых представляет переходную группу от диподасковых к высшим аскомицетам (образование сумки по способу крючка, примитивной аскогенной гифы), с одной стороны, и к своеобразной группе грибов, вегетативное тело которых состоит из одиночных клеток (сумчатым дрожжам), с другой стороны.
Семейство Сахаромицетовые (Sacchabomycetaceae) (И. П. Бабьева)
Представители семейства сахаромицетовых (Saccharomycetaceae) не образуют типичного мицелия, их вегетативные клетки почкуются или делятся. Аскосиоры образуются в сумках, представляющих одиночные клетки. У многих дрожжей в цикле развития удлиняется диплоидная фаза, а у некоторых она преобладает.
Под общим названием дрожжи с этими грибами часто объединяют почкующиеся или делящиеся грибы, не образующие сумчатой стадии (аспорогенные дрожжи). Некоторые из таких несовершенных дрожжей имеют родственные связи с базидиальными грибами.
Дрожжами называют грибы, которые существуют на протяжении всего или большей части жизненного цикла в виде раздельных одиночных клеток. Дрожжи обладают всеми основными свойствами и признаками грибных организмов, являясь органотрофными эукариотами с абсорбционным типом питания, но одноклеточное их строение влечет за собой ряд последствий. Одноклеточные организмы имеют более высокую скорость обмена веществ (в расчете на единицу массы), чем мицелиальные грибы, благодаря большему значению отношения площади поверхности к объему.
Дрожжи растут и размножаются с большой скоростью, вызывая при этом существенные изменения в окружающей среде. Наиболее известный для дрожжей процесс спиртового брожения был причиной широкого практического использования дрожжей с давних времен. Именно это их свойство привело к тому, что дрожжи исторически всегда рассматривают отдельно от других грибов. Автономность группы дрожжей в научном отношении поддерживается также тем, что методы их изучения более сходны с бактериологическими, чем микологическими. Однако дрожжи сохраняют до сих пор самостоятельность среди других грибов скорее лишь по традиции, чем по общебиологическому принципу. Постоянное обнаружение все большего числа новых переходных и промежуточных форм среди мицелиальных грибов и дрожжей делает все более трудным и условным проведение границ между теми и другими.
Филогенетические связи обнаруживаются у дрожжей как с сумчатыми (аскомицетами), так и с базидиальными грибами. Подразделение дрожжей на группы, размещающиеся в разных классах грибов (Ascomycetes или Basidiomycetes), базируется главным образом на способах их полового размножения. Дрожжи, у которых половой цикл не обнаружен, временно относят к классу несовершенных грибов (Fungi Imperfecti, или Deuteromycetes).
В современной грибной таксономии имеется тенденция объединять все дрожжевые и дрожжеподобные организмы в один особый класс Endomycetes, включающий грибы, у которых гаплоидная фаза представлена почкующимися клетками, плодовые тела отсутствуют, а половой процесс соматогамного типа (копуляция соматических клеток, а не специализированных гамет). Образующаяся в результате соматогамии зигота или непосредственно превращается в спорангий (сравните с сумкой аскомицетов), или прорастает дикарионтическим мицелием, на котором образуются телиоспоры, дающие начало промицелию со споридиями (сравните с базидией базидиомицетов).
Таким образом, термин "дрожжи" не имеет номенклатурной ценности и не признан в ботанической таксономии. Термины, которыми обозначают дрожжи в разных языках, так или иначе связаны с явлениями или процессами, сопровождающими брожение. Французское levure происходит от латинского levere, означающего подъем, т. е. вспенивание жидкости при брожении за счет выделения углекислого газа. Немецкое Hefe имеет в основе глагол heben - поднимать, а английское yeast и родственное голландское gist происходят от греческого zestos - кипение, пена. В русском языке термин "дрожжи" имеет корень, общий со словами "дрожь", "дрожать", которые применимы в описании вспенивающейся бродящей жидкости.
Общность происхождения терминов, отражающих функции дрожжевых организмов и процессы брожения, свидетельствует о том, что человек издавна связывал их между собой. По-видимому, бродильные процессы были первыми на пути использования человеком деятельности микроскопических существ. Можно считать, что дрожжи - это самые древние из культивируемых человеком растений.
Применение дрожжей для приготовления алкогольных напитков из соков фруктов и ягод, из зерен злаков (после их осахаривания) уходит корнями в доисторические времена. Археологические раскопки в Египте и Двуречье обнаруживают хорошо сохранившиеся остатки пивоварен и хлебопекарен, которые были построены за 2000 и даже 6000 лет до н. э. Известно, что в древнем Вавилоне была развита техника получения солода и пивоварение, ассирийцы умели готовить вино еще за 3500 лет до н. э. Легенды древних греков говорят о том, что искусство приготовления вина было даровано людям богом Дионисом.
Первый открыл причинную связь между брожением и дрожжами основоположник микробиологии Л. Пастер, обобщивший свои взгляды и наблюдения в знаменитой книге "Etudes sur la biere" ("Очерки о пиве"), опубликованной в 1876 г. Л. Пастер установил, что брожение, являясь энергетическим процессом, заменяет дыхание тем организмам, которые живут в анаэробных условиях: "La fermentation est vie sans air" ("Брожение есть жизнь без воздуха"). Еще за 10 лет до этого (1866) Пастер опубликовал "Исследования о вине, его болезнях и причинах, которые их вызывают. Новые способы сохранения и старения вина". На русском языке этот труд впервые полностью был опубликован в 1960 г. в двухтомном издании Академии наук СССР (Луи Пастер. Избранные труды). В нем имеются иллюстрации, на которых изображены дрожжевые клетки, а также дано описание способов и аппаратов для стерилизации вина. Предложенный Пастером метод стерилизации для сохранения вина путем прогрева при 50-60 °С получил название пастеризации - приема, который широко применяется сейчас в разных отраслях пищевой промышленности.
Как живые существа дрожжи были описаны задолго до открытия Пастера. Первым увидел их голландец Антони ван Левенгук. Рисунки дрожжевых клеток с описаниями он направил в 1680 г. в Королевское общество в Лондон. Эту дату связывают с открытием мира микроорганизмов.
Значение факта обнаружения Левенгуком дрожжей не было понято ни его современниками, ни учеными последующих поколений в течение 150 лет. Научно-ботанические описания дрожжей, их строения и размножения были сделаны только в 30-х годах прошлого столетия почти одновременно К. Латуром во Франции (1838) и Т. Шванном (1837, 1839) и Ф. Кютцингом (1837) в Германии. В результате этих работ дрожжи были отнесены к грибам. Хотя уже первые исследователи дрожжей видели в них структуры, которые были ими определены как споры, на роль спор в размножении обратили внимание значительно позже де Сенез (1868) и Рисс (1869- 1870).
Рисс назвал спорообразующие дрожжи "сахарными грибами" (Saccharomyces). Это название, ранее использованное Мейеном (1837) для почкующихся дрожжей, сохраняется до сих пор за одним из самых известных и хорошо изученных родов дрожжевых организмов, к которым относятся все так называемые "культурные" дрожжи (винные, пивные, пекарские).
Морфолого-систематические исследования дрожжей, описания многих новых видов, выделенных из природных источников, и разработка первых классификационных схем связаны с именем датского ботаника Э. X. Хансена, работавшего на протяжении 30 лет в конце прошлого и начале нашего столетия. Работами Хансена не только заложено начало совершенно новой эры в биологии дрожжей, но и капитально перестроена технология брожения. Никто из его предшественников, в том числе и Пастер, не работал с чистыми культурами. Хансен разработал метод получения чистых культур из одной клетки и показал, что отдельные штаммы сильно различаются по физиологическим свойствам. Только внедрение чистых культур дало возможность получать пиво устойчивого качества, а пивоварение после этого стало развиваться как широкая промышленная отрасль.
Серия работ по систематике дрожжей выполнена так называемой голландской (делфтской) школой микробиологов, выпустивших в свет несколько изданий определителей дрожжевых организмов.
На русском языке (и в немецком переводе) имеется хорошо известная специалистам монография В. И. Кудрявцева "Систематика дрожжей" (1954).
В конце 60-х - начале 70-х годов таксономисты разных стран объединяют усилия для проведения общей классификации дрожжей, соответствующей современному уровню знаний об этих организмах. В результате появляется в Амстердаме издание книги "Дрожжи" (1970) под общей редакцией Дж. Лоддер с участием 13 авторов. В этой книге описано свыше 360 видов дрожжей и 39 родов.
Широкие цитологические исследования дрожжей, начатые французским ученым А. Гийермоном в начале столетия, активно развивались в Советском Союзе Г. А. Надсоном и его учениками. Новая область цитологических исследований была открыта благодаря разработке и внедрению методов люминесцентной микроскопии М. Н. Мейселем.
Если начало физиологическому направлению в изучении дрожжей положили работы Л. Пастера, то первым фактом, явившимся основой для биохимических исследований, следует считать открытие братьями Бюхнер в 1897 г. зимазного комплекса. Они пытались приготовить экстракт из пивных дрожжей для медицинских целей и для этого растирали дрожжи с трепелом, а затем на механическом прессе отжимали сок. Добавив к бесклеточному соку в качестве консерванта сахар, они обнаружили с удивлением, что сок начал бродить. Фактор, ответственный за бесклеточное брожение, был назван зимазой и состоял, как теперь известно, из смеси энзимов. Слово "энзим" происходит из греческого языка и означает "в дрожжах". Открытие зимазного комплекса - это начальная веха в развитии биохимии. Вскрытие механизма спиртового брожения показало, что аналогичные реакции гликолиза лежат в основе того процесса, благодаря которому любой живой организм получает из сахара энергию для жизни.
Вклад дрожжей в развитие биологических наук огромен. Дрожжи - прекрасная модель для изучения многих процессов и явлений. На дрожжевых объектах выполнены первые исследования по радиобиологии (Г. А. Надсон и Г. С. Филиппов), сделаны цитологические и генетические открытия (цитоплазматическая наследственность, генетическая самостоятельность митохондрий и др.), имеющие общебиологическое значение.
Промышленное использование дрожжей базируется прежде всего на их способности превращать сахар в спирт и углекислоту, а также на их воздействии на зерновые и молочные продукты. Главную роль дрожжи играют в производстве пива, вина, сидра; в получений путем дистилляции крепких спиртных напитков (водка, виски, бренди, коньяк, джин, ликеры); в хлебопекарной промышленности. В последние годы к старым технологическим процессам добавились новые: производство промышлен-ного спирта из отходов целлюлозно-бумажной промышленности или мелассы; получение дрожжевой биомассы или дрожжевых экстрактов для использования в качестве кормовых или пищевых добавок; применение дрожжей как фармацевтического продукта для лечебных целей (главным образом из-за высокого содержания витаминов группы В); получение из дрожжей или с помощью дрожжей различных ценных биохимикатов, например эргостерина (витамина D2), липидов, нуклеиновых кислот, ферментов и коферментов, органических кислот.
Для характеристики масштабов производства и использования дрожжей можно привести в качестве примера тот факт, что одних только пекарских дрожжей ежегодно в мире производят не менее 700 000 т, а сухих кормовых дрожжей - приблизительно 200 000 т. В будущем производство дрожжей будет еще расширяться, особенно в связи с обнаружением новых источников сырья для дрожжевых производств (например, углеводороды нефти).
Итак, дрожжи всегда, сколько помнит себя человечество, его неизменные спутники и друзья. Они помогают человеку и сегодня. Еще больше можно ждать от них в будущем, так как знания наши об этих удивительных организмах еще далеко не полные.
Дрожжи в культурах на плотных питательных средах растут в виде колоний разного цвета, формы и консистенции (табл. 13), а в жидких средах образуют муть, пленки и осадки. Колонии дрожжей на первый взгляд мало отличаются от бактериальных: они не опушены воздушным мицелием как у актиномицетов и грибов и чаще всего бывают гладкими, густыми и плотными или реже - слизистыми, растекающимися. По цвету они могут быть чисто-белыми, буровато-бежевыми, коричневыми (при спорообразовании) или яркими, окрашенными во все тона желто-оранжево-красного цвета. Описан-ные так называемые "черные дрожжи" не являются на самом деле дрожжами, а представляют собой дрожжевидные стадии разных темноцветных грибов.
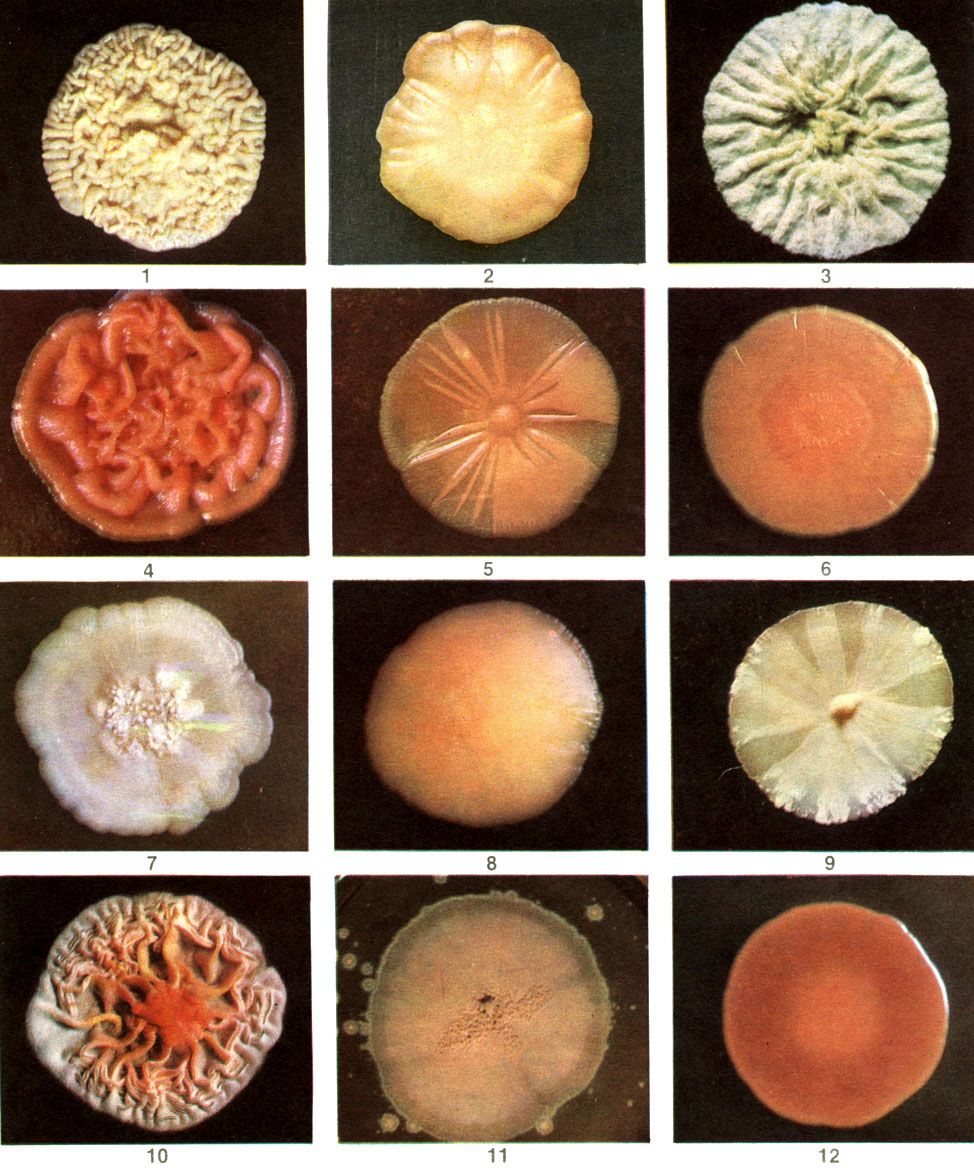
Таблица 13. Колонии дрожжевых грибов на питательной среде: 1 - пекарские дрожжи (Saccharomyces cerevisiae); 2 - мечниковия прекраснейшая (Metschnikowia pulcherrima); 3 - кандида земляная (Candida humicola); 4 - родоторула клейкая (Rhodotorula glutinis); 5 - родоторула красная (R. rubra); 6 - родоторула золотистая (R. aurantiaca); 7 - дебариомицес Кантарелли (Debaryomyces cantarelli); 8 - криптококк Лавра (Cryptococcus laurentii); 9 - надсония продолговатая (Nadsonia elongata); 10 - спороболомицес розовый (Sporobolomyces roseus); 11 - спороболомицес хольсатикус (S. holsaticus); 12 - родоспоридиум диобоватум (Rhodosporidium diobovatum)
Таллом дрожжей в наиболее простой форме представляет одну клетку. Эта клетка имеет все основные структуры, характерные для грибов (рис. 54). Иногда клетки могут объединяться в различные более или менее прочные структуры в виде ложного или настоящего мицелия. Способ организации клеток в такие структуры может быть характерным для некоторых видов и родов дрожжей (табл. 14).
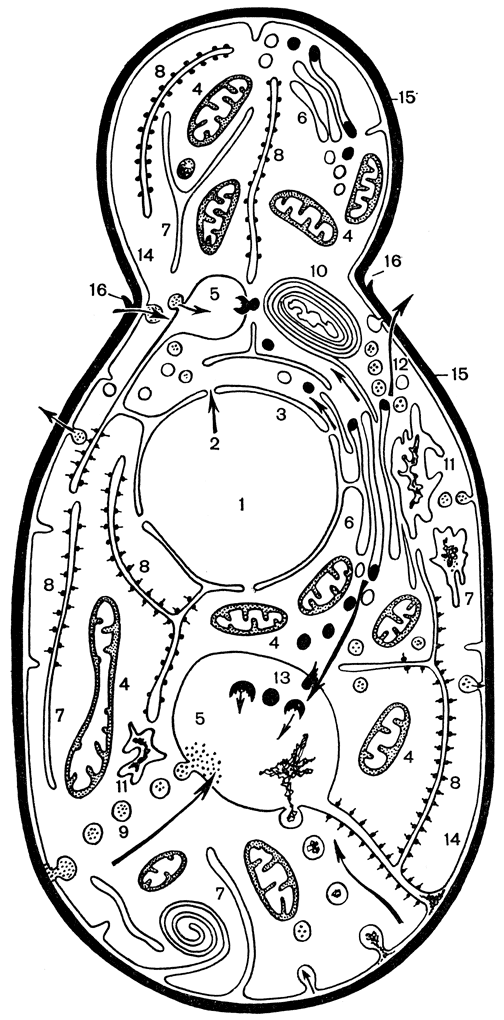
Рис. 54. Схема строения дрожжевой клетки: 1 - ядро с ядерной порой (2) и мембраной (3); 4 - митохондрии; 5 - вакуоля; 6 - мембраны комплекса Гольджи; 7 и 8 - гладкий и шероховатый эндоплазматический ретикулум; 9 - пиноцитозные пузырьки; 10 - сегрегационные гранулы; 11 - фагосомы; 12 - выделительные пузырьки; 13 - липидные включения; 14 - цитоплазматическая мембрана; 15 - клеточная стенка; 16 - кольцевой валик шрама почкования
Таблица 14. Ультра структура дрожжевой клетки
Размеры одиночных дрожжевых клеток лежат в пределах от 1 до 10 мкм, чаще 3-7 мкм. При образовании мицелия или вытянутых клеток псевдомицелия длина их может достигать 20 и даже 50 мкм, в то время как ширина остается более постоянной и обычно не превышает 10 мкм.
Форма дрожжевой клетки в значительной степени зависит от способа вегетативного размножения. Оно может происходить путем почкования, деления и почкующегося деления. При почковании возникают круглые, яйцевидные или овальные клетки (рис. 55,1). Почкование - наиболее характерный и широко распространенный у дрожжей способ вегетативного размножения. Начинается почкование с появления на поверхности клетки небольшого сферического выроста, который увеличивается в размерах и как бы "отшнуровывается" от материнской клетки, оставляя на ней шрам, или почечный рубец.
Одна клетка может иметь до 3-4 десятков почечных рубцов, по числу дочерних клеток. На вновь образовавшейся молодой клетке сохраняется шрам рождения, менее заметный, чем почечный рубец. Первая почка на новой клетке появляется, как правило, на противоположном полюсе от шрама рождения. Почки у многих видов и родов дрожжей возникают неупорядоченно, т. е. на любой части поверхности клетки, по одной или по две и более одновременно. Такой тип почкования принято называть многосторонним, а в последнем случае - множественным (рис. 51,2). Иногда почки закладываются по полюсам, или на широком основании, но затем почкование заканчивается обычным образом (рис. 55,3,4). Среди почкующихся дрожжей есть такие, которые выделяются в самостоятельные роды из-за особой формы их клеток. Это дрожжи с треугольными (Trigonopsis ), серповидными (Selenotila) или стреловидными (Brettanomyces) клетками (рис. 55,5-7). В результате почкования без отделения клеток у некоторых дрожжей возникают структуры примитивного или хорошо развитого ложного мицелия с бластоспорами и бластоконидиями (рис. 55,9). Последнее характерно для рода кандида (Candida).
Почкующееся деление отличается от истинного почкования тем, что почка, закладывающаяся на широком основании и обычно у одного из полюсов, после созревания не "отшнуровывается", а "откалывается" по образующейся в перешейке поперечной перегородке (рис. 55,10). Клетки при этом приобретают грушевидную, веретеновидную или лимоновидную форму.
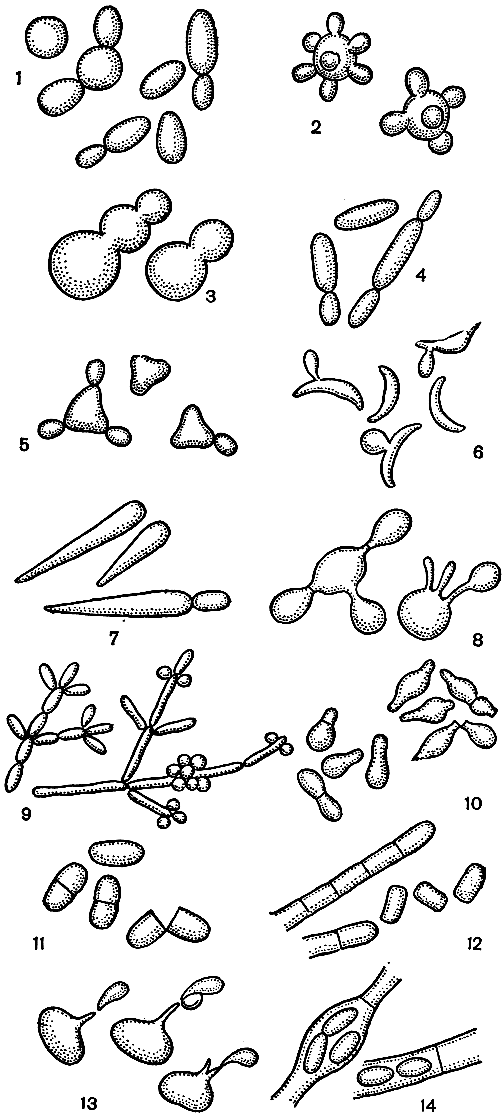
Рис. 55. Способы вегетативного размножения дрожжей (Объяснение в тексте.)
Деление распространено среди дрожжей значительно реже, чем почкование. Клетки после деления могут оставаться одиночными, как у шизосахаромицетов (Schizosaccharomyces, рис. 55,11), или образуют мицелий, который иногда распадается на артроспоры, как у трихоспорона (Trichosporon, рис. 55,12). В последнем случае клетки располагаются в виде характерного зигзага, напоминающего вагоны поезда, сошедшего с рельсов.
Необычный способ вегетативного размножения путем почкования на длинных выростах описан у дрожжей, которые выделены на этом основании в самостоятельный род стеригматомицес (Sterigmatomyces, рис. 55,8).
Образование с силой отстреливающихся клеток - баллистоспор, сидящих на концах заостренных стеригм, характерно для спороболомицетов (Sporobolomyces, рис. 55,13) и буллеры (Bullera). У некоторых дрожжей описаны вегетативные эндоспоры, которые возникают в отдельных клетках или в клетках мицелия в количестве от 2 до 10 и более (рис. 55,14).
Дрожжи могут существовать в гаплоидной и диплоидной фазах. Смена ядерных фаз рассматривается как чередование поколений у дрожжей. Оно связано с образованием половых спор, которое может происходить по аскомицетному или базидиомицетному типу.
К группе дрожжей, имеющих аскомицетный цикл развития, относят больше половины известных родов дрожжевых организмов.
У аскомицетных дрожжей в результате копуляции после плазмо- и кариогамии зигота непосредственно развивается в сумку, где диплоидное ядро после первого деления в мейозе претерпевает редукцию. Образующиеся аскоспоры всегда гаплоидны. Однако у некоторых дрожжей наблюдается своеобразное извращение в чередовании поколений: прорастающие споры или первые получившиеся из них гаплоидные клетки сливаются попарно и размножаются дальше почкованием уже в диплоидном состоянии. У таких дрожжей сумка со спорами образуется без непосредственно предшествующей копуляции.
В зависимости от продолжительности существования гапло- и диплофазы различают гаплоидные и диплоидные дрожжи. Жизненные циклы тех и других приведены на рисунке 56. В качестве примеров диплоидных дрожжейг которые длительно вегетативно размножаются, находясь в диплофазе, выбраны сахаромикоды Людвига (Saccharomycodes ludwigii), гансениаспора апикулятная (Hanseniaspora apiculata) и пекарские дрожжи (Saccharomyces cerevisiae). Различия в их половом цикле проявляются в разных способах восстановления диплофазы после образования гаплоидных спор. Это могут быть копуляция прорастающих спор (S'codes ludwigii, рис. 56,1), диплоидизация ядра в одной споре (H'spora apiculata, рис. 56,2), копуляция образовавшихся из спор мелких гаплоидных клеток или одной клетки и споры (S. cerevisiae, рис. 56,3). Гаплоидные дрожжи в природе длительно существуют и вегетативно размножаются в гаплофазе. Диплоидизация перед образованием спор происходит по-разному: копуляция двух отдельных клеток (Schizosaccharomyces, рис. 56,4), слияние ядер материнской и дочерней клеток в так называемой мейозной почке (Debaryomyces, Schwanniomyces, рис. 56,5), кратковременное образование дикариона, миграция его в специальную почку, из которой после кариогамии и мейоза формируется сумка со спорой (Nadsonia, рис. 56,6).
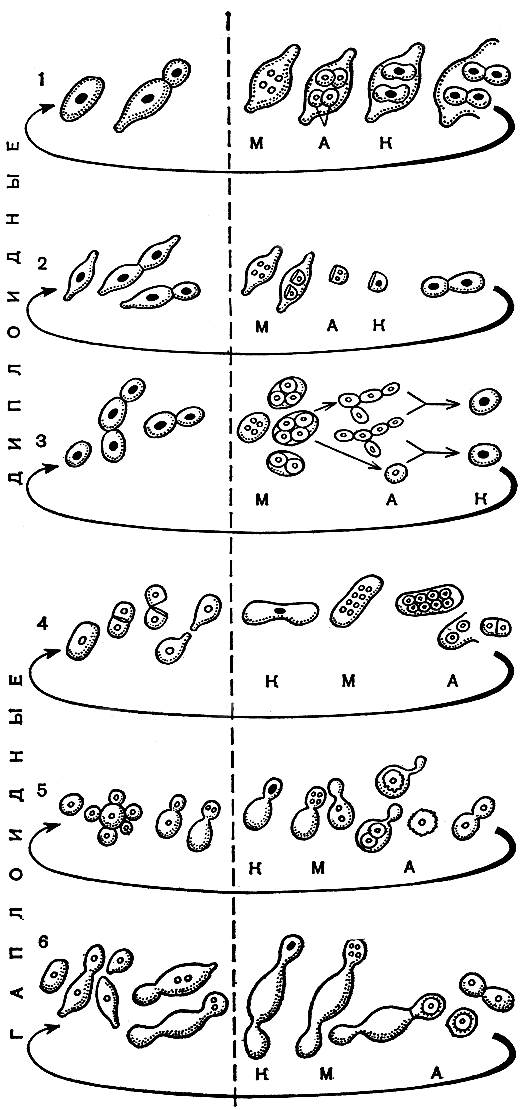
Рис. 56. Жизненные циклы и чередование ядерных фаз у гаплоидных и диплоидных дрожжей: к - кариогамия; м - мейоз; а - аскоспоры. (Объяснение в тексте.)
Имеются и другие формы полового процесса, заканчивающиеся также образованием сумок со спорами. У дрожжей рода Metschnikowia сумки образуются из хламидоспор, в которые превращаются крупные диплоидные клетки после предшествующей копуляции мелких гаплоидных клеток противоположных типов спаривания (рис. 57). Сумки в виде крупных мешковидных выростов с многочисленными спорами образуются у почвенных липомицетов (Lipomyces).
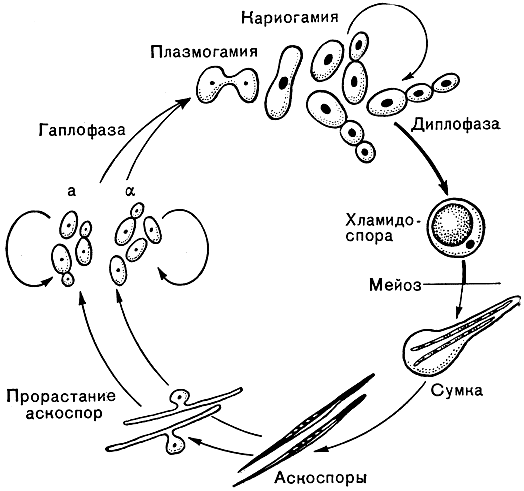
Рис. 57. Цикл развития гриба мечниковия прекраснейшая (Metschnikowia pulcherrima)
У дрожжей известна раздельнополость (гетероталлизм). Именно это явление - причина того, что многие дрожжи известны только в несовершенной стадии и не обнаруживают полового процесса. Такие дрожжи рассматриваются в классе несовершенных грибов. Однако сейчас для некоторых видов подобраны типы спаривания и, таким образом, найдены их совершенные аналоги. В некоторых родах (Hansenula, Pichia) объединяются гомо- и гетероталличные виды.
Смеси клеток разных типов спаривания часто, хотя и не всегда спорулируют более активно, а при смешивании они могут агглютинировать (склеиваться), образуя плотный осадок на дне сосуда.
Половая агглютинация может, иметь и практическое значение. На основе этого явления разработан метод быстрого и эффективного выделения дрожжей из жидкости в производстве пекарских дрожжей, кормовых дрожжей, в пивоварении, дистилляции и виноделии.
Эндогенные аскоспоры, образующиеся в результате полового процесса, различаются по форме и содержатся в разном количестве в сумках.
На электронных микрографиях, а иногда даже в световом микроскопе можно видеть на поверхности спор различные скульптурные образования типа выростов, бородавок, ободков или складок (рис. 58).

Рис. 58. Аскоспоры дрожжей: 1 - сахаромицеты; 2 - клюйверомицеты; 3 - дебариомицеты; 4 - шванниомицеты; 5 - надсония; 6-7 - липомицеты; 8 - викерхамия; 9 - мечниковия; 10 - пихия; 11 и 12 - эндомикопсис; 13 - сахаромикоды
Помимо дрожжей, у которых половой процесс приводит к образованию споровых вместилищ с эндогенными спорами, существует большая группа дрожжевых организмов с экзогенным образованием половых спор. Эти споры (споридии) формируются на проростках, называемых иногда промицелием. Такие проростки возникают из особых телиоспор (телейтоспор) после некоторого периода покоя. Жизненный цикл этих дрожжей имеет много общего с циклом развития головневых грибов (порядок Ustilaginales).
Мысль о родстве некоторых дрожжевых организмов с базидиальными грибами была высказана впервые в 1924 г. Клюйвером и ван Нилем при описании ими баллистоспоровых дрожжей. Эти дрожжи получили родовое название спороболомицес (Sporobolomyces) на том основании, что они образуют отстреливающиеся с силой баллистоспоры. В основе отстреливания спор лежит капельно-экскреторный механизм, как и у многих базидиомицетов.
Аналогичные баллистоспоры были найдены и у следующего, описанного в 1930 г. Дерксом, рода дрожжей Буллера (Bullera). Однако полового цикла ни у того, ни у другого рода долгое время не находили, а их предполагаемое родство с базидиомицетами основывали лишь на общности образования и отстреливания баллистоспор.
Прямое доказательство существования у дрожжей базидиомицетного жизненного цикла было продемонстрировано впервые в 1967 г. японским исследователем Банно на "красных" дрожжах родоторула (Rhodotorula). Сложный половой цикл у родоторулы клейкой (R. glutinis) включает конъюгацию клеток разных типов спаривания с объединением ядер в дикарионы, прорастание образующейся зиготы дикариофитным мицелием с пряжками, формирование телиоспоры, в которой происходит кариогамия, сопровождающаяся мейозом в процессе прорастания промицелием и образованием гаплоидных клеток-споридий (рис. 59). Дрожжи с новым половым циклом были выделены в самостоятельный род, родоспоридиум (Rhodosporidium). Аналогичный половой процесс обнаружен в настоящее время у многих других дрожжей, в том числе и у некоторых видов несовершенных родов - кандида (Candida) и спороболомицес (Sporobolomyces).
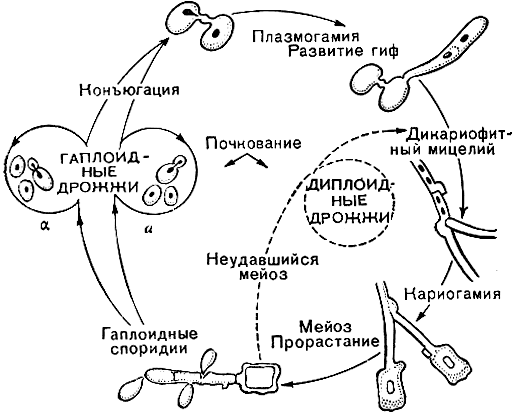
Рис. 59. Схема жизненного цикла красных дрожжей родоторула-родоспоридиум (Rhodotorula-Rhodosporidium)
Наиболее известный и характерный для дрожжей процесс обмена веществ - спиртовое брожение. Однако те же самые дрожжи, которые вызывают этот процесс (пекарские, пивные, винные), обладают также резко выраженным аэробным (дыхательным) обменом веществ. Оба процесса являются энергетическими, хотя последний более экономичен и дает значительно больший выход энергии. Дыхательный путь обмена веществ у дрожжей включает цикл трикарбоновых кислот, глиоксилатный и пентозный циклы. Каждый из них характеризуется накоплением в среде специфических продуктов обмена: разных органических кислот или многоатомных спиртов. Побочными продуктами могут быть необычные вещества класса липидов: гликолипиды, сфинголипиды. Многие организмы, включаемые в настоящее время в группу дрожжей, совсем не обладают способностью к брожению, а проводят только прямое окисление субстрата (например, Rhodotorula, Sporobolomyces, Cryptococcus, Lipomyces и др.).
В качестве источников углерода дрожжи используют лучше всего различные сахара, а также простые и многоатомные спирты, органические кислоты и многие другие соединения. Неизвестны дрожжи, которые не потребляли бы глюкозу. Некоторые виды используют углеводороды парафинового ряда, циклические углеводороды и сложные соединения ароматического строения (фенол, нафталин). В использовании дрожжами разных источников питания находят отражение условия их природных местообитаний. Разрушение дрожжами сложных природных и неприродных соединений типа пектина, крахмала, ароматических веществ имеет отношение к превращению органических соединений в природе. Эти процессы могут быть важными также в связи с проблемой очистки сточных вод от промышленных предприятий.
Ценными во многих отношениях особенностями дрожжей являются, с одной стороны, их потребность в витаминах и, с другой, способность к сверхсинтезу некоторых из них. Дрожжи крайне различаются своими потребностями в витаминах. Факторами роста для дрожжей служат витамины группы В. Те штаммы, которые характеризуются абсолютной потребностью в одном или двух витаминах, могут быть использованы в качестве индикаторов на присутствие этих витаминов в субстрате и для количественного их определения. Такие методы для шести витаминов группы В - биотина, тиамина, инозита, пиридоксина, пантотеновой и никотиновой кислот - с использованием дрожжевых тест-объектов разработаны в нашей стране Е. Н. Одинцовой.
Дрожжи-сверхсинтетики используются для промышленного получения некоторых витаминов. С помощью специальных рас дрожжей производят также обогащение витаминами и микроэлементами некоторых продуктов питания (например, молочнокислые продукты, дрожжевые галеты).
Необычными потребностями для роста характеризуются дрожжи, обитающие в специфических природных очагах. Сюда относятся, например, постоянные обитатели кишечника теплокровных животных. Некоторые из них обладают очень высокими потребностями в витаминах и аминокислотах (Saccharomycopsis guttulata из кишечника кролика), все они могут расти только при повышенных температурах, в диапазоне 27-45° С (например, Candida slooffii из кишечника свиней).
Дрожжи, выделяемые из арктических и антарктических вод и из почв полярных районов, проявляют, наоборот, свойство психрофилии: они не растут при температуре выше 20° С. Облигатно-психрофильными видами являются, например, дрожжи рода лейкоспоридиум (Leucosporidium).
К осмофильным дрожжам, которые лучше растут при высоких концентрациях сахара в среде, относятся, например, сахаромицес меллис (Saccharomyces mellis) и сахаромицес Роукса (S. rouxii). Их находят в меде и других средах с большим содержанием сахара.
Среди дрожжей нет галофильных (солелюбивых) видов, а есть лишь галотолерантные, т. е. устойчивые к большой концентрации солей, но не требующие высокого содержания соли в среде.
Среди дрожжей де Бари (Debaryomyces), выделяющихся часто из морской воды, из рассолов, из солонины, есть виды, которые хотя и медленно, но могут расти в почти насыщенных растворах поваренной соли.
Среди дрожжей нет видов, образующих токсические для человека вещества. При порче пищевых продуктов, вызываемой дрожжами, меняется вкус и внешний вид, но не происходит накопления вреднодействующих веществ, как это бывает у ядовитых грибов и бактерий.
Но есть виды дрожжей, патогенные для растений и человека, причиняющие существенный вред. Патогенность дрожжей по отношению к растениям проявляется значительно слабее, чем у бактерий и некоторых фитопатогенных грибов. Дрожжи с игловидными спорами Nematospora coryli поражают главным образом южные растения: хлопчатник, некоторые виды южной фасоли, цитрусовые. Переносчиками этих дрожжей, вероятно, служат насекомые. Есть сведения о клопах как носителях и переносчиках возбудителей заболевания хлопчатника - болезни сердцевины семенной коробочки. Cryptococcus neoformans - опасный возбудитель болезни мозга человека и животных, которая зачастую кончается фатально. По имени возбудителя болезнь называется криптококкозом. В природе эти дрожжи часто обнаруживаются в помете голубей, которые, возможно, являются переносчиками возбудителя.
Другой патогенный для человека вид дрожжей - Candida albicans - поражает наружные покровы и внутренние органы, вызывая глубокие системные микозы - кандидозы, болезни слизистых и сепсис. Благоприятными условиями для проявления болезни является общее истощение организма или же искусственные дисбактериозы под действием неправильного применения антибиотических препаратов.
Говоря о разных взаимоотношениях дрожжей с высшими и низшими организмами, о роли дрожжей в жизни других существ, следует упомянуть об их пищевом значении для некоторых простейших и насекомых. В кишечнике плодовой мушки дрозофилы постоянно находятся дрожжи, главным образом те виды, которые обитают на поверхности плодов и ягод. Откладывая яйца в мякоть плодов, мушка одновременно вводит туда и дрожжи. Находясь в благоприятной среде, дрожжи активно размножаются и образуют значительную биомассу, которой питаются вылупляющиеся из яиц личинки. Взрослые мушки, перелетая с одного субстрата на другой, переносят и распространяют дрожжи.
Особая группа дрожжевых организмов, главным образом из родов пихия (Pichia) и гансепула (Hansenula), ассоциирована с насекомыми-ксилофагами, например с жуками-короедами, поражающими хвойные деревья. Некоторые из этих насекомых гибнут в стерильных условиях, лишенные дрожжей-симбионтов. Дрожжи из этих местообитаний используют в качестве питательных веществ целлобиозу и ксилозу - продукты расщепления гемицеллюлоз древесины. Они устойчивы к растительным дубильным веществам и способны развиваться на средах с некоторыми танинами в качестве источника углерода.
Дрожжи служат хорошей пищей для амеб. Почвенные амебы прекрасно размножаются на дрожжевом корме, поедая живые и мертвые клетки разных видов дрожжей.
Как уже было сказано выше, дрожжи объединяются в отдельную группу на основании весьма поверхностного сходства и главным образом на основании морфологических признаков. Исследование тонких механизмов их половой репродукции, биохимических процессов, лежащих в основе обмена веществ и химии отдельных структур, приводит к выводу о большой разнородности этой группы организмов и, следовательно, разных путях происхождения и эволюции отдельных семейств и родов дрожжей.
Порядок эндомицетов объединяет два семейства аскоспоровых дрожжей. В семействе спермофторовых (Spermophthoraceae) все дрожжи имеют игловидные споры, а семейство сахаромицетных (Saccharomycetaceae) объединяет все остальные дрожжи с разной (но не игловидной) формой спор. В это семейство входят те роды, которые имеют наибольшее хозяйственное значение и изучение которых положило начало развитию наших знаний о дрожжевых организмах.
Род дробянки, или шизосахаромицеты (Schizosaccharomyces), объединяет дрожжи, вегетативно размножающиеся только делением. Это гаплоидные дрожжи, клетки которых копулируют перед образованием сумок. В сумке обычно 4 или 8 спор. Споры круглые, овальные или бобовидные, гладкие или с бородавчатой поверхностью. У большинства дробянок споры дают положительную йодную реакцию, что свидетельствует о наличии в их оболочках крахмалоподобных соединений.
Наиболее изученные виды - шизосахаромицет помбе (Sch. pombe) и шизосахаромицет восъмиспоровый (Sch. octosporus) (рис. 60). Морфологически эти виды различаются размерами клеток (у второго вида они более крупные - до 18 мкм в длину и 8 мкм в ширину) и числом спор в сумках: 4 у первого и 8 - у второго.
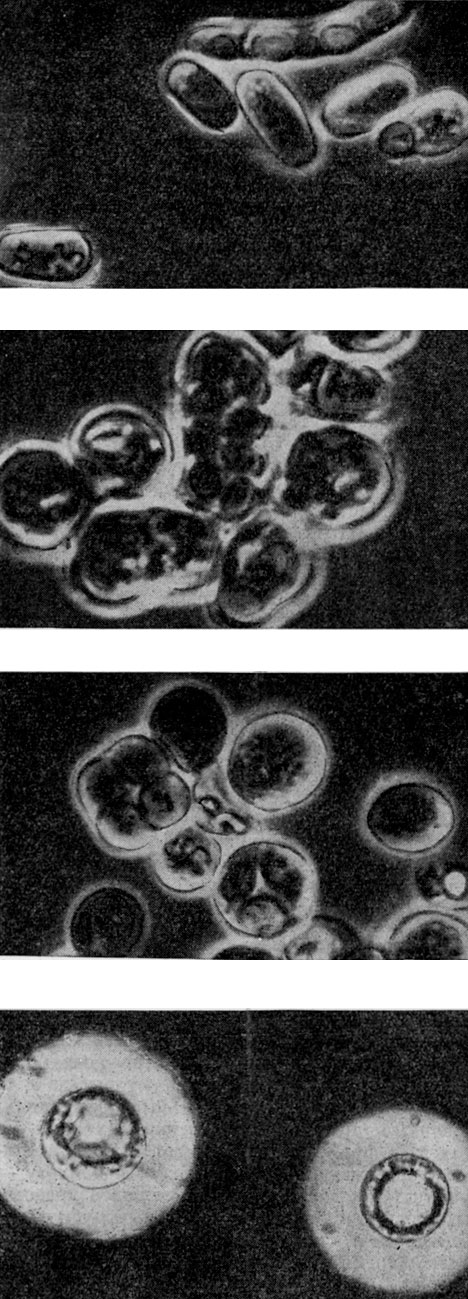
Рис. 60. Микрофотографии дрожжей (сверху вниз): Schizosaccharomyces pombe (вегетативные клетки и сумка с четырьмя спорами); Schizosaccharomyces octosporus (делящиеся клетки и сумка с восемью спорами); Saccharomyces cerevisiae (вегетативные клетки и сумка со спорами); Lipomyces starkeyi (капсулы)
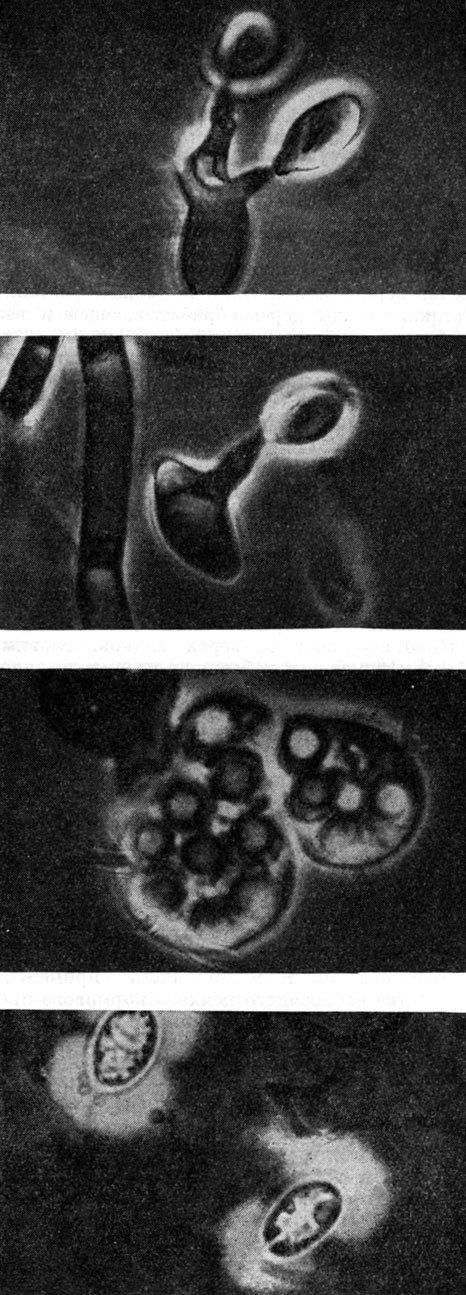
Рис. 61. Микрофотографии дрожжей (сверху вниз): Sporobolomyees roseus (клетки со стеригмами и баллистоспорами); то же; Lipomyces starkeyi (сумка со спорами); Cryptococcus diffluens (капсулы)
Шизосахаромицеты - энергичные возбудители спиртового брожения. Шизосахаромицет помбе, получивший свое видовое имя по названию африканского пива "помбе", используют во многих тропических странах для приготовления особых сортов пива и крепких алкогольных напитков (рома, аррака). Распространение этих дрожжей в странах жаркого климата объясняется тем, что эти грибы хорошо растут только при повышенных температурах (30- 42° С). Из природных субстратов вне хозяйства человека эти дрожжи почти не выделяются. Можно думать поэтому, что шизосахаромицет помбе - "культурные" дрожжи, выведенные человеком в жарких странах в процессе изготовления спиртных напитков аналогично производственным расам сахаромицетов, которые созданы в странах умеренного климата.
Род сахарные грибы, или сахаромицеты (Saccharomyces), объединяет как природные виды, так и "культурные" (производственные), существование которых тесно связано со всей историей бродильной промышленности. Наиболее общим признаком всех дрожжей этого рода является их способность к активному сбраживанию Сахаров с образованием наибольшего, по сравнению с другими дрожжами, количества спирта (от 10 до 19% по объему). Сюда входят пивные, винные и пекарские дрожжи.
Морфологически все виды этого рода довольно сходны: они имеют сферические и овальные клетки, размножающиеся вегетативно почкованием. Все они образуют овальные и шаровидные аскоспоры в сумках, возникающих либо после непосредственно предшествующей копуляции гаплоидных клеток, либо из диплоидных клеток без предварительной копуляции. Число спор в сумках обычно не больше четырех (рис. 60).
В литературе было описано более 200 видов сахаромицетов, однако сейчас различают не более 20 из них. Наибольшее значение для человека имеет S. cerevisiae. Этот вид известен в сотнях рас "культурных" дрожжей.
Производственные дрожжи разделяют на верховые (винные, хлебопекарные, спиртовые расы) и низовые (пивные).
Вина с помощью дрожжей получают из виноградных и плодово-ягодных соков. Вина бывают сухие, сладкие и крепленые. В первых сахар сброжен полностью, во вторых - не до конца, а в третьи добавляют спирт. При производстве игристых вин (шампанское) брожение завершают в герметически закрытых бутылках, где идет накопление углекислоты. Получение разных вин зависит от технологии процесса, исходного сырья и свойств применяемых рас дрожжей.
Винодельческие расы получают свои названия в зависимости от страны и местности, из которых они произошли, а также от сорта винограда и местоположения виноградника. На основе практически важных свойств дрожжей расы делят на группы. Например, известны высоковыбраживающие расы, дающие до 18 и 20,5% спирта; холодостойкие или, наоборот, термотолерантные расы; спиртоустойчивые расы для шампанских вин; так называемые хересные дрожжи, образующие на поверхности вина пленку и создающие характерный для хереса вкус и букет. Последние иногда рассматривают не как расу S. cerevisiae, а как отдельный вид (хересный вариант) S. oviformis var. cheresiensis (В. И. Кудрявцев 1954).
Сырьем для получения спирта биологическим способом, кроме пищевых продуктов (картофель, зерновые), служат кормовые растения, патока, отходы деревообрабатывающей и целлюлозной промышленности (сульфитные щелока). Так как спиртовые дрожжи не используют сложные сахара (полисахариды), содержащиеся в этих продуктах, то их подвергают предварительному осахариванию (гидролизу) солодом или кислотами. Солод - это проросший ячмень, содержащий ферменты амилазы, которые расщепляют крахмал. Дрожжи затем сбраживают образующиеся моносахара в спирт. В производстве этилового спирта получаются также другие ценные побочные продукты, например фурфурол, метиловый спирт, углекислота. Спиртовое брожение можно направить и в сторону накопления глицерина.
Пиво получают из зерен злаков, главным образом ячменя, и добавляют на определенной стадии женские соцветия (шишки) хмеля. Полученное охмеленное сусло сбраживают дрожжами. Конечный продукт брожения (пиво) содержит до 6% спирта, сахар, экстрактивные вещества, белки, органические кислоты, горькие вещества хмеля, дубильные вещества и углекислоту. Дрожжи, применяемые в пивоварении, относят к видам S. carlsbergensis и S. uvarum. Первый из них иногда рассматривают как расу или разновидность пекарских дрожжей.
Дрожжи, которые используют в разных бродильных производствах, накапливаются в качестве отходов и могут быть применены в качестве добавочного ценного кормового продукта в пищевые рационы животных. Эти дрожжи содержат белки, углеводы, жиры и минеральные соли.
Дрожжевая биомасса хорошо усваивается также организмом человека. Поэтому дрожжи специально получают и для лечебных целей. Их применяют в жидком виде и в таблетках. 500 г сухих дрожжей заменяет по белку 1 кг свежего мяса, 33 штуки куриных яиц или 4,1 л коровьего молока. В качестве витаминных препаратов достаточна суточная доза в 25 г сухих или 100 г прессованных дрожжей. Перед употреблением их следует убить прогреванием (растереть с водой и залить кипятком) и добавлять в различные блюда.
Пекарские дрожжи (S.cerevisiae) применяют в хлебопекарной и кондитерской промышленности. Для этих целей специально производят прессованные дрожжи, введенные в практику хлебопечения в конце XIX в. Дрожжи размножают в условиях высокой аэрации. Чисто спиртовое брожение под действием дрожжей происходит в пшеничном тесте. Образующаяся при брожении углекислота вызывает подъем теста. В ржаном тесте идет смешанное брожение: сначала молочнокислое (бактериальное), а затем спиртовое (дрожжевое). Из-за образующейся в первом этапе молочной кислоты ржаной хлеб имеет кисловатый вкус.
Среди сахаромицетов есть и природные виды, распространенные главным образом в субстратах, содержащих сахар: на поверхности плодов, ягод и фруктов, в нектаре цветов, в сокотечениях деревьев. Некоторые виды ассоциированы с насекомыми и встречаются в местах их обитания. Так называемые осмофильные дрожжи, известные как S. rouxii, обитают в пчелином меде. Эти дрожжи лучше используют фруктозу (сахар меда), чем глюкозу. Раньше гаплоидные осмофильные дрожжи выделяли в особый род зигосахаромицетов (Zygosaccharomyces). Осмофильные дрожжи часто являются причиной порчи меда, варений, джемов, а также скисания вин.
К роду сахаромицетов близки как активные возбудители брожения дрожжи рода клюйверомицетов (Kluyveromyces), названные по имени голландского микробиолога Клюйвера. В этот род перенесены некоторые виды, которые раньше рассматривались как сахаромицеты или зигосахаромицеты. Это дрожжи с бобовидными спорами (K. marxianiis, K. fragilis), сбраживающие молочный сахар - лактозу. Они постоянно выделяются из молока и молочнокислых продуктов, в образовании которых им принадлежит существенная роль. Некоторые виды этого рода (K. polysporus, K. africanus) образуют многоспоровые сумки, и выделяют их из почв в Южной Африке.
Дрожжи близких между собой родов пихия (Pichia) и гансенула (Hansenula) дифференцируются в настоящее время главным образом на основании их отношения к нитратам, а именно по способности восстанавливать нитраты в процессе ассимиляции. Первый род относится к нитратотрицательным, а второй - к нитратположительным дрожжам. Провести четкую границу между видами той и другой групп не всегда легко. Среди этих родов есть как гаплоидные, так и диплоидные виды, гомо- и гетероталличные. Споры у тех и других чаще всего шляпообразные вследствие сдвинутого к полюсу ободка. Есть также сатурновидные и просто круглые с шероховатой поверхностью споры. У большинства видов споры легко освобождаются из сумок после созревания. Гаплоидные штаммы, образующие псевдомицелий, могут быть приняты за представителей рода кандида (Candida), внутри которого описаны аспорогенные аналоги пихия и гансенула. Физиологически виды описываемых родов относятся к группе слабо бродящих дрожжей, а некоторые (особенно среди пихий) совсем не вызывают брожения. На поверхности жидких питательных сред или вина эти дрожжи образуют пленки, поэтому в виноделии их относят к группе облигатных пленочных дрожжей.
Особенно заражаются пленочными дрожжами вина с низким содержанием спирта, находящиеся в условиях хорошей аэрации. Дрожжи окисляют спирт и остаточные сахара вина, а в среде накапливаются органические кислоты, альдегиды и эфиры. Таким образом, пленочные дрожжи родов пихия и гансенула являются представителями вредной микрофлоры брожения. Меры предохранения и борьбы с ними предусматривают специальные режимы подготовки и сбраживания перерабатываемого сырья. Решающее значение имеет ведение процесса брожения на чистых расах сильных (активно бродящих) культурных дрожжей.
Некоторые пихии и гансенулы - это свободноживущие дрожжи, другие ассоциированы с насекомыми, особенно с жуками-короедами хвойных деревьев. Такие виды, как сатурновая, ароматная и калифорнийская гансенулы (H. saturnus, H. suaveolens u H. californica), часто выделяются из почв, особенно из луговых и болотных. Служит ли для них почва постоянным природным местообитанием, где они размножаются, или же эти дрожжи заносятся в почву из других очагов, еще не совсем ясно.
Род дебариомицеты (Debaryomyces), названный по имени знаменитого ботаника А. де Бари, объединяет дрожжи с характерным половым процессом: перед образованием сумки происходит копуляция материнской клетки с клеткопочкой. Таким образом, дрожжи эти в природе гаплоидны. Споры, чаще всего по одной в сумке, имеют бородавчатую или с правильными выростами поверхность (рис. 58). Клетки дебариомицетов чаще круглые, реже яйцевидные и овальные. Виды со сферическими клетками размножаются вегетативно множественным почкованием (рис. 55). Среди дебариомицетов есть дрожжи небродящие, слабо сбраживающие сахара, и только дрожжи двух или трех видов активно сбраживают разные углеводы.
Дебариомицеты отличаются от других дрожжей тем, что многие из них хорошо развиваются на белковых средах и при повышенных концентрациях солей в среде. Дебариомицет Ганзена (D. hansenii) часто выделяется из морской воды. Дебариомицеты постоянно обнаруживают на поверхности копченых колбас, сыров, сосисок. Это - сухой белый налет или тонкая белая пленочка на поверхности рассолов при засоле капусты, огурцов. Дебариомицеты часто выявляются при микозных заболеваниях кожи человека и животных, хотя их патогенность остается недоказанной. Недавно было установлено, что два вида - дебариомицет Кантарелли (D. cantarellii) и дебариомицет муравьиный (D. formicarius) - связаны с рыжими лесными муравьями группы формика руфа (Formica rufa L.), в гнездах которых они развиваются в количестве, превышающем иногда 10 млн. клеток на 1 г субстрата.
Близки к дебариомицетам по циклу развития шванниомицеты (Schwanniomyces, по имени Т. Шванна). Все описанные 4 вида этого рода пока выделены только из почв. Споры их имеют характерную для этого рода форму грецкого ореха (рис. 58).
Особняком среди почкующихся аскоспоровых дрожжей стоят жировые дрожжи - липомицеты (Lipomyces). Как и шванниомицеты, липомицеты распространены только в почвах. Эти дрожжи можно считать типичными представителями почвенной дрожжевой флоры. Клетки этих дрожжей крупные (до 9-10 мкм в диаметре), чаще сферические, реже слегка овальные. Размножаются липомицеты многосторонним почкованием, причем почки иногда закладываются на широком основании. Клетки их содержат много жира (отсюда название рода) и окружены слизистой полисахаридной капсулой (рис. 60). По типу образования спор все липомицеты разбиваются на две группы, четырехспоровые и многоспоровые. Споры тех и других различаются по структуре поверхности: ребристая у четырехспоровых и складчатая или гладкая у многоспоровых (см. рис. 58). Способ образования сумок не совсем ясен, однако показано,что это дрожжи гомоталличные.
Все виды липомицетов не вызывают брожения и относятся к олигонитрофилам, т. е. группе микроорганизмов (чаще всего почвенных обитателей), которые нормально развиваются при следовых количествах азота в среде. В качестве источников углерода они способны окислять многие сахара, в том числе и пентозы, а также многоатомные спирты.
На территории Советского Союза липомицеты были найдены в почвах, начиная от подзолов в зоне северной тайги и кончая каштановыми почвами зоны сухих степей. Они выделялись также из некоторых почв Крыма, Кавказа и Тянь-Шаня.
Аскоспоровые дрожжи с почкующимся делением, выделяемые иногда в семейство сахаромикодовых (Saccharomycodaceae), распределяются в 4 рода: надсония (Nadsonia), сахаромикоды (Saccharomycodes), гансепиаспора (Hanseniaspora) и викерхамия (Wickerhamia). Эти дрожжи имеют лимоновидные клетки: мелкие у гансениаспоры и очень крупные у всех остальных (например, у сахаромикодов до 34 мкм в длину). Дрожжи этих родов различаются по форме спор, способу диплоидизации и образования сумок, по природным местообитаниям. У сахаромикодов Людвига (S. ludwigii) споры круглые, с узким гребешком. В сумке обычно 4 споры, расположены они тесно, попарно, гребешками друг к другу. При прорастании эти споры парами конъюгируют, будучи еще в сумке, и дают диплоидные клетки. Наиболее часто эти дрожжи обнаруживаются в слизетечении дуба, а также на поверхности плодов и в забродивших плодово-ягодных соках. В виноделии этот вид относят к сорнякам брожения.
Находят сахаромикоды Людвига и в так называемом "чайном квасе", где они обитают в симбиозе с уксуснокислыми бактериями.
Этот слабокислый напиток широко распространен в Индонезии. Для его приготовления используют настой чая с добавлением 10% сахара. На поверхности жидкости "чайный гриб" образует мощную слизистую пленку. Дрожжи, сбраживая сахар, способствуют образованию небольшого количества спирта и углекислого газа.
Серьезными вредителями в промышленном виноделии являются дрожжи гансепиаспора (Hanseniaspora), известные в специальной литературе под названием апикулятусов. Они широко распространены на ягодах винограда, на мягких фруктах. Дрожжи гансениаспоры вызывают энергичное брожение, а образуемые при этом продукты задерживают развитие винных дрожжей. Накапливающиеся летучие кислоты и эфиры придают вину нежелательные вкус и запах. Основной метод борьбы с апикулятусами - сульфитирование плодово-ягодных соков. Для гансениаспоры известны аспорогенные аналоги, которые описывают как виды рода клёкера (Kloeckera). Они чаще встречаются в районах северного виноделия.
Дрожжи Надсона (Nadsonia) - N. fulvescens и N. elongata - имеют желто-коричневые споры с хорошо заметными даже в светооптическом микроскопе выростами (рис. 58). Спорулирующие культуры обычно темно-коричневого цвета. Эти дрожжи - слабые бродилыцики. В природе они постоянно встречаются в весенних эксудатах деревьев в зоне умеренного климата. Последнее объясняется низким температурным оптимумом этих дрожжей: выше +25° С они не растут. Дрожжи Надсона впервые были выделены и описаны Г. A. Hадсоном (совместно с А. Г. Конокотиной) в 1911 г.
Последний род из этой группы - викерхамия (Wickerhamia) - представлен единственным видом - викерхамия флюоресцирующая (W. fluorescens), видовой эпитет которому дан по выделяемому флюоресцирующему пигменту - рибофлавину. Эти дрожжи образуют характерные кепковидные с козырьком споры (рис. 58), чаще по одной в сумке. Найдены они пока только в помете дикой белки в Японии.
Из семейства спермофторовых (Spermophthoraceae), образующих игловидные споры, наиболее изучен род дрожжей Мечникова (Metschnikowia). Он объединяет несколько видов, которые четко делятся на две группы по их местообитанию - водные и наземные. Впервые представители этого рода были обнаружены П. И. Мечниковым (1884) в теле дафний из бассейна Парижского ботанического сада.
Дафнии заглатывают с водой дрожжевые сумки со спорами. Под действием желудочного сока оболочки сумок растворяются, а игловидные споры прокалывают стенку кишечника и выходят в полость тела, где прорастают и размножаются почкованием. При этом иногда животное гибнет, дрожжи переходят в состояние спор и вновь попадают в окружающую среду, откуда идет заражение новых экземпляров дафний. Более 80 лет об этих дрожжах дополнительно ничего не было известно. И только в 60-х годах, когда развернулись широкие микробиологические исследования океана, были найдены в теле разных беспозвоночных аналогичные дрожжи, описанные как новые виды: M. krissii и M. zobellii, названные в честь советского микробиолога и исследователя океанов А. Е. Крисса, а также американского водного микробиолога К. Зобелла.
В настоящее время в этом роде описаны еще два вида дрожжей, местообитаниями которых служат наземные субстраты, в первую очередь растения. Это мечниковия прекраснейшая и мечниковия Реукауфа (M. pulcherrima u M. reukaufii), известные ранее как нектарные дрожжи (Nectaromyces) или как аспорогенные дрожжи кандида (Candida pulcherrima).
Дрожжи этих видов обнаруживают в нектаре цветов (чаще M. reukaufii) или на поверхности листьев, плодов и фруктов (M. pulcherrima). Возможно, переносчиками их являются насекомые-опылители. Хотя жизненный цикл этих дрожжей в природе еще полностью не изучен, но по аналогии с водными обитателями можно предположить, что часть этого цикла (возможно, связанная с образованием спор) проходит в теле насекомых-переносчиков. Развитие этих дрожжей в чистых культурах в лабораторных условиях показано на рисунке 57. М. pulcherrima легко отличается от других дрожжей тем, что образует внеклеточный пигмент пульхерримин, окрашивающий среду (в присутствии железа) в кирпично-красный цвет.
Баллистоспоровые дрожжи рода спороболомицеты (Sporobolomyces) размножаются бесполым путем почкованием и отстреливающимися баллистоспорами почковидной формы (рис. 61). Колонии этих дрожжей имеют от бледно-розовой до ярко-красной окраски за счет синтезируемых каротиноидных пигментов (табл. 14). При посеве штрихом в чашку Петри дрожжи образуют на крышке зеркальное отражение этого штриха благодаря отстреливанию баллистоспор (рис. 60). В природе эти дрожжи обитают в составе эпифитной микрофлоры и особенно распространены на листовой поверхности культурных и диких злаков. С листьев дрожжи переходят на семена, где и перезимовывают, вновь заражая растения весной при прорастании семян. Их вредное действие на растения не доказано, они являются типичными сапрофитами, хотя численность их значительно увеличивается на слабых (увядающих или пораженных болезнью) растениях.
Несовершенные дрожжевые организмы, размножающиеся только вегетативным путем и не образующие баллистоспор, составляют разнородную группу, члены которой не имеют между собой близкого родства, а многие представители являются просто аспорогенными двойниками споровых дрожжей (все Kloeckera, многие из Torulopsis, Candida, Brettanomyces). В этой группе имеются роды дрожжей, которые размножаются почкованием (например, Torulopsis), почкующимся делением (Kloeckera, Schizoblastosporion), делением (Trichosporon) и образованием новых клеток на отростках, напоминающих иногда стеригмы (Sterigmatomyces, Sympod iomyces).
Широко распространены в природе красные дрожжи рода родоторула (Bhodotorula). Их находят на поверхности листьев растений, в слизетечениях деревьев, воздухе, почве, в пресных и морских водоемах. Наиболее характерным их природным местообитанием является, по-видимому, листовая поверхность многих растений, где численность этих дрожжей в летний период достигает многих сотен тысяч клеток на 1 г субстрата. Выше указано, что для одного из видов - родоторула клейкая (Bh. glutinis) - в настоящее время описан совершенный цикл, который имеет много общего с жизненным циклом головневых грибов (см. рис. 59).
Имеются единичные указания на случаи заболевания растений под действием этих дрожжей. Однако чаще всего красные дрожжи ведут сапрофитный образ жизни, развиваясь как безвредные эпифиты на поверхности листьев.
Дрожжи-криптококки (Cryptococcus), подобно красным дрожжам, можно обнаружить в самых разнообразных местах, как в природе, так и в хозяйстве человека. Отдельные виды, однако, имеют более определенные местообитания и ограниченные ареалы распространения. Больше всего работ посвящено патогенному виду Cr. neoformans. Клетки этих дрожжей круглые, многосторонне почкующиеся, мицелия и псевдомицелия не образуют. С поверхности они одеты слизистой капсулой (рис. 61). В состав внеклеточных полисахаридов криптококков входит крахмал. Колонии имеют слизистую консистенцию и часто стекают по штриху на дно пробирки.
Вид Cr. laurentii, слабо образующий каротиноидные пигменты, от которых колонии в старых культурах приобретают розоватый или желтоватый оттенок (табл. 14), чаще всего обнаруживается среди эпифитных микроорганизмов на разных растениях, как древесных, так и травянистых. Такие виды криптококков, как белый (Cr. albidus), почвенный (Cr. terreus) и терриколюс (Cr. terricolus), постоянно обитают в почвах. Последний вид рассматривается как перспективный продуцент липидов.
Роды торулопсис (Torulopsis) и кандида (Candida) различаются между собой тем, что дрожжи кандида способны формировать мицелий и псевдомицелий. Дрожжи обоих родов имеют очень широкие границы и включают разнообразные виды, которые из-за отсутствия специфических характеристик не могут быть помещены в другой род аспорогенных дрожжей.
Торулопсис - бесцветные почкующиеся дрожжи, обладающие слабой (большинство видов) или активной бродильной способностью. Некоторые образуют капсулы, но во внеклеточных полисахаридах, в отличие от криптококков, нет крахмалоподобных веществ. Многие описанные виды являются гаплоидными штаммами аскоспоровых сахаромицетов, дебариомицетов и клюйверомицетов. Выделяются эти дрожжи из природных источников и из очагов, связанных с деятельностью человека. Специфические субстраты и местообитания отдельных видов исследованы мало.
Все дрожжи рода кандида, помимо почкования, образуют псевдомицелий, а иногда и истинный мицелий, но не артроспоры в отличие от трихоспорона (Trichosporon). Псевдомицелий варьирует от примитивного до хорошо развитого, с бластоспорами. Такие дрожжи рода кандида, как С. albicans и С. stellatoidea, образуют характерные хламидоспоры, прорастающие вегетативными клетками. Основываясь на этом признаке, некоторые исследователи (ван дер Вальт, фон Аркc) сближают эти виды с базидиомицетами. Другие виды кандида, наоборот, известны как аспорогенные варианты аскомицетных дрожжей родов пихия, гансенула, эндомикопсис и клюйверомицес. Большая группа психрофильных кандид (С. scottii, С. nivalis, С. gelida, С. frigida) перенесена во вновь описанный род дрожжей - лейкоспоридиум (Leucosporidium) - с базидиомицетным жизненным циклом. При описании рода мечниковия упоминалось о том, что два вида, включенные в него, ранее были известны как виды рода кандида. Таким образом, ясно, что кандида - это один из самых неопределенных и гетерогенных в систематическом отношении родов дрожжей.
Особое внимание в последние годы кандида привлекли к себе тем, что дрожжи некоторых видов (С. guilliermondii, С. tropicalis, С. lipolytica) хорошо растут на углеводородах, синтезируя при этом много белка, липидов и других ценных веществ.
Кормовые дрожжи (С. utilis) тоже относятся к этому роду. Их белки содержат все необходимые животным аминокислоты. Получают кормовые дрожжи из разного сырья: мелассы, кислотных гидролизатов древесины, сульфитных щелоков и отходов сельскохозяйственного производства. С. utilis - неприхотливые дрожжи: они используют, кроме гексоз, также пентозные сахара и не нуждаются в добавках витаминов для роста.
Семейство Спермофторовые (Spermophthoraceае) (И. И. Сидорова)
Семейство спермофторовых объединяет роды, образующие настоящий мицелий (Nematospora, Eremothecium и Spermophthora), и роды с почкующимися клетками (Metschnikowia, см. выше). Для этого семейства характерны удлиненные, веретеновидные, часто заостренные аскоспоры, образующиеся в сумках от 1 до 32. Грибы этой группы - паразиты растений, развивающиеся преимущественно на плодах, возбудители так называемых стигматомикозов. Инфекция распространяется насекомыми, и грибы проникают в растения через сделанные ими проколы.
Большинство видов этой группы обитает в районах с теплым климатом, часто в субтропиках. Некоторые из них имеют практическое значение как возбудители болезней культурных растений (например, нематоспора лещиновая - возбудитель гнили фисташки). Эремотециум Эшби - сверхсинтетик витамина В2 (рибофлавина) - используется для его промышленного получения.
У хлопчатниковой спермофторы (Spermophthora gossypii) - паразита в коробочках хлопчатника - в цикле развития чередуются гаплоидное и диплоидное поколения, различающиеся морфологически (гетероморфная смена поколений). Это один из немногих известных у грибов случаев смены поколений в цикле развития. Она отсутствует у всех высших грибов (аскомицетов и базидиомицетов) и известна еще только у хитридиомицетов в порядках хитридиевых (Physoderma) и бластокладиевых (Blastocladiella и Allomyces).
Гаплоидный мицелий гриба лишен перегородок либо немногочисленные перегородки делят его на крупные многоядерные клетки. Этот мицелий может образовывать почкующиеся клетки. На нем интеркалярно (вставочно по ходу гифы) образуются удлиненные спорангии, содержащие многочисленные (до 40) веретеновидные споры (рис. 62). Эти споры могут прорастать в новый гаплоидный мицелий, т. е. функционировать как споры бесполого размножения, но чаще они функционируют как гаметы, попарно копулируя и образуя зиготу. Из зиготы развивается ветвящийся диплоидный мицелий, состоящий из одноядерных клеток. На концах его гиф образуются шаровидные сумки, содержащие по восемь гаплоидных аскоспор.
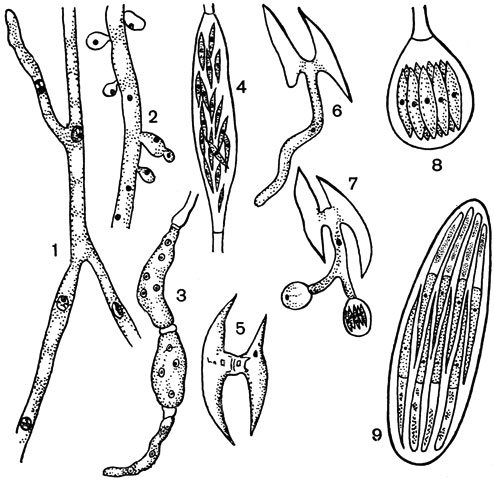
Рис. 62. Спермофторовые. Спермофтора хлопчатника (Spermophthora gossypii) (1-8): 1 - гаплоидный мицелий; 2 - блаетоспоры на гаплоидном мицелии; 3-4 - развитие спорангиев; 5 - слияние спор; 6-7 - прорастание зиготы; 8 - сумка. Нематоспора фасолевая (Nematospora phaseoli): 9 - сумка
Прорастая, аскоспоры снова дают начало гаплоидному неклеточному мицелию (рис. 62).
Эремотециум Эшби (Eremothecium ashbyi), также паразитирующий на коробочках хлопчатника, образует дихотомически ветвящийся мицелий ярко-желтого цвета, состоящий из многоядерных клеток. Окраска мицелия гриба обусловлена присутствием рибофлавина, который накапливается в нем в таких количествах, что выпадает в виде кристаллов в вакуолях. На мицелии образуются продолговатые многоспоровые спорангии с веретеновидными спорами. Оболочка спорангия после созревания лопается, споры освобождаются и прорастают не сливаясь, в отличие от предыдущего вида. На образовавшихся проростках развиваются сумки, содержащие от 4 до 32 аскоспор (рис. 63).
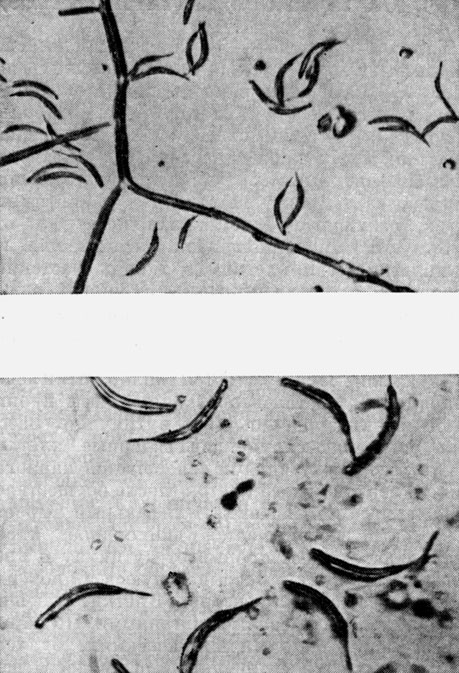
Рис. 63. Аскоспоры эремотециума Эшби (Eremothecium ashbyi)
Гриб хорошо растет на питательных средах и используется в микробиологической промышленности для получения рибофлавина. Для производства витамина эремотециум выращивают в глубинной культуре при хорошей аэрации на средах с органическими источниками азота, витаминами и углеводами при температуре 28- 30 °С. В этих условиях мицелий гриба синтезирует большое количество рибофлавина, выделяющегося в среду. Его выход составляет 1-2 г, а по некоторым данным - до 6 г чистого вещества на 1 л среды.
В животноводстве используют концентраты рибофлавина или обогащенные им добавки к кормам. Для их получения продуцент выращивают на зерне или различных отходах пищевой промышленности. Субстрат при выращивании на нем гриба сохраняет свои пищевые качества и обогащается витамином, содержание которого доходит до 2-3 г на 1 кг сырья.
Для рода нематоспора (Nematospora) характерны восьмиспоровые сумки и веретеновидные двуклеточные аскоспоры с длинным придатком на одном конце (рис. 62). Мицелий у видов этого рода может почковаться.
Виды рода нематоспора паразитируют на различных растениях, вызывая гниль семян и мясистых частей плодов. Лещиновая нематоспора (N. coryli) вызывает гниль лесных орехов, а в последние годы обнаружена на фисташке в ряде стран Северной' Африки и Азии, где причиняет большой ущерб. В республиках Средней Азии в 1967-1970 гг. она вызвала потери урожая фисташки до 30% (Н. В. Кутлунина). Гриб поражает семя, которое сначала темнеет, а затем сгнивает, покрываясь белым налетом аскоспор и почкующихся клеток гриба. Околоплодник не поражается, и орехи внешне выглядят здоровыми. Только расколов их, можно обнаружить признаки заболевания.
В культуре этот вид образует хорошо развитый мицелий и почкующиеся клетки. Диплоидизация происходит в результате слияния клеток мицелия или аскоспор, и зигота развивается в сумку сразу, или сначала из нее вырастают гифы, а на них образуются сумки. Смены поколений нет.
Известны также нематоспоры, паразитирующие на других растениях. На хлопчатнике в некоторых районах Африки, Азии, Северной и Центральной Америки встречается хлопчатниковая нематоспора (N. gossypii), на фасоли и других растениях - фасолевая нематоспора (N. phaseoli), а на томатах - томатовая нематоспора (N. lycopersici).
Спермофторовые представляют интересную группу низших аскомицетов, сочетающую примитивные признаки предков (неклеточный гаплоидный мицелий, образование спорангиев, смена поколений в цикле развития) с продвинутыми признаками аскомицетов (клеточный мицелий, образование сумок с фиксированным числом аскоспор). По увеличению продолжительности диплоидной фазы в цикле развития спермофторовые сходны с некоторыми сахаромицетами.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'