Класс сумчатые грибы, или аскомицеты (Ascomycetes) (И. И. Сидорова)
Сумчатые грибы, или аскомицеты,- один из крупнейших классов грибов. В нем более 30 000 видов, что составляет около 30% всех известных видов грибов. Входящие в этот класс грибы чрезвычайно разнообразны по строению. Сюда относятся, например, дрожжи, представленные одиночными почкующимися клетками, и виды с плодовыми телами различной формы и размеров, от микроскопических до крупных, достигающих иногда 10-20 см (сморчки, строчки). Но все эти многообразные формы связаны общим происхождением и имеют ряд общих черт, на основании которых они объединяются в этот класс.
Основной признак аскомицетов - образование в результате полового процесса сумок (или асков) - одноклеточных структур, содержащих фиксированное число аскоспор, обычно 8 (рис. 50). Сумки образуются или непосредственно из зиготы (у низших аскомицетов), или на развивающихся из зиготы аскогенных гифах. В сумке происходит слияние ядер зиготы, а затем мейотическое деление диплоидного ядра и образование гаплоидных аскоспор. У высших аскомицетов сумка представляет не только место образования аскоспор, но и активно участвует в их распространении.
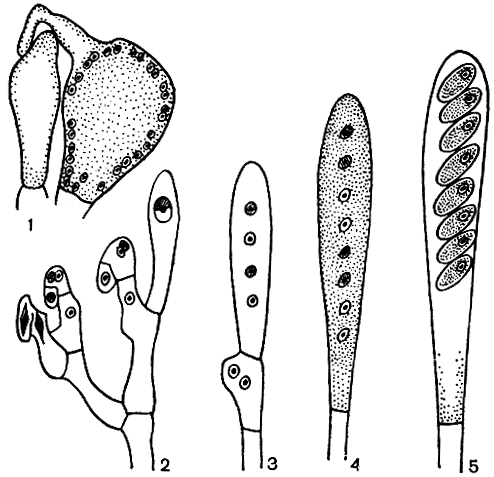
Рис. 50. Половой процесс и развитие сумок у аскомицетов: 1 - аскогон с трихогиной и антеридий; 2 - развитие сумок по способу крючка; 3 - молодая сумка после мейоза; 4 - молодая сумка с восемью гаплоидными ядрами; 5 - зрелая сумка с аскоспорами
Вегетативное тело аскомицетов - разветвленный гаплоидный мицелий, состоящий из многоядерных или одноядерных клеток. В отличие от зигомицетов перегородки (септы) в мицелии аскомицетов образуются упорядоченно, синхронно с делением ядер. Развитие септ происходит от стенок гифы к центру, напоминая сужение диафрагмы в объективе фотоаппарата. В центре септы остается пора (рис. 51), через которую происходит движение цитоплазмы с со скоростью от 1-2 до 25-40 см/ч. Некоторые органеллы клетки, даже ядра, могут мигрировать через поры. Наличие пор в септах играет существенную роль в переносе питательных веществ по гифам в зону роста.
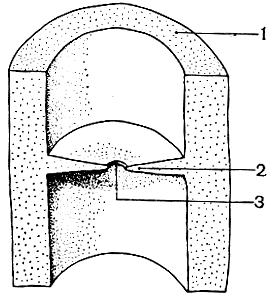
Рис. 51. Септа (перегородка) в мицелии аскомицета: 1 - наружная стенка гифы; 2 - септа; 3 - пора
У некоторых аскомицетов мицелий может распадаться на отдельные клетки или почковаться. У дрожжей (порядок Endomycetales) настоящего мицелия нет, а вегетативное тело представлено одиночными почкующимися, реже делящимися клетками, иногда образующими псевдомицелий. Дрожжеподобный рост наблюдается и у некоторых мицелиальных аскомицетов, например у грибов-дерматофитов (порядок Eurotiales), тафриновых (порядок Taphrinales), некоторых видов из рода цератостис (порядок Microascales).
Высокоспециализированные экзопаразиты насекомых из порядка лабульбениевых (Laboulbeniales) имеют редуцированный мицелий, а их тело (рецептакл) состоит из настоящей ткани.
В состав клеточных стенок аскомицетов, как и у хитридиомицетов и зигомицетов, входит хитин, но его содержание у грибов этого класса ниже и составляет не более 20-25% полисахаридов клеточной стенки (для сравнения: у некоторых хитридиомицетов - до 60%, у зигомицетов - до 37-40%). У некоторых дрожжей (род Schizosaccharomyces) хитин не обнаружен. Большую часть полисахаридов клеточной стенки аскомицетов (80-90%) составляют глюканы - полимеры D-глюкозы, отличающиеся от целлюлозы характером связи между мономерами. У дрожжей, кроме глюканов, обнаружены маннаны - полимеры маннозы. Целлюлоза у аскомицетов не обнаружена, за исключением двух видов из рода цератоцистис.
В цикле развития аскомицетов большую роль играет бесполое размножение. Споры бесполого размножения (конидии) образуются на гаплоидном мицелии экзогенно (реже эндогенно) на конидиеносцах различного строения. Конидиальные спороношения аскомицетов очень разнообразны по морфологии. Конидиеносцы образуются одиночно на мицелии, соединяются в пучки (коремии) или подушечки (спородохии), развиваются плотным слоем на поверхности сплетения гиф (ложа) или внутри шаровидных либо грушевидных споровместилищ с отверстием на вершине (пикниды). Типы конидиальных спороношений и различные способы образования конидий подробно описаны в главе о дейтеромицетах, или несовершенных грибах.
Конидиальные спороношения развиваются в период вегетации грибов и служат для их массового расселения. У аскомицетов - паразитов растений они обычно образуются на живом растении, а сумчатые спороношения, за немногими исключениями,- после отмирания растения или его частей, в конце периода вегетации или после перезимовки.
У некоторых аскомицетов конидиальное спороношение неизвестно, у других оно преобладает в цикле развития. В отдельных группах этого класса наблюдается редукция полового процесса и сумчатая стадия образуется редко. Иногда ее трудно обнаружить в природе и получить в искусственной культуре грибов. Поэтому многие аскомицеты как в природе, так и в коллекциях культур мы чаще встречаем в их конидиальной стадии. Это отражается и на их названиях. Конидиальные стадии многих аскомицетов имеют самостоятельные видовые наименования, и в литературе мы чаще встречаем их именно под этими названиями, а не под названиями сумчатых стадий. Например, широко используемый в генетических исследованиях аскомицет Emericella nidulans более известен как Aspergillus nidulans. Распространенный в почве и на различных целлюлозосодержащих субстратах аскомицет Нуросгеа rufa обычно называют Trichoderma viride по его конидиальной стадии. Особенно распространено это для фитопатогенных аскомицетов: название Fusariumsolani вместо Nectriahaematococca, Helminthosporium sativum вместо Cochliobolus sativus и т. д. В Международном кодексе ботанической номенклатуры, который регулирует употребление названий растений, содержится даже специальный пункт, разрешающий использовать наряду с основным видовым названием аскомицета (по его сумчатой стадии) название его конидиальной стадии в тех случаях, когда речь идет именно об этой стадии гриба. Это единственный случай, когда для разных стадий в цикле развития одного организма допускаются самостоятельные видовые названия.
Для большого числа грибов, встречающихся в природе в гаплоидной конидиальной стадии, половые стадии неизвестны. Такие грибы относят к классу дейтеромицетов, или несовершенных грибов.
Половой процесс, типичный для аскомицетов,- гаметангиогамия, т. е. слияние двух гаметангиев - специализированных клеток, не дифференцированных на гаметы.
У низших аскомицетов (подкласс Hemiascomycetidae) половой процесс сходен с зигогамией у зигомицетов. Гаметангии разного пола морфологически сходны или малоразличимы и представляют выросты или веточки мицелия. После их слияния сразу происходит кариогамия и сумка развивается непосредственно из зиготы. Однако, в отличие от зигомицетов, в многоядерных гаметангиях сливаются только два ядра (нет множественной кариогамии), зигота не переходит в состояние покоя, а сразу развивается в сумку. В цикле развития низших аскомицетов, следовательно, есть только гаплоидная и диплоидная стадии (рис. 52).
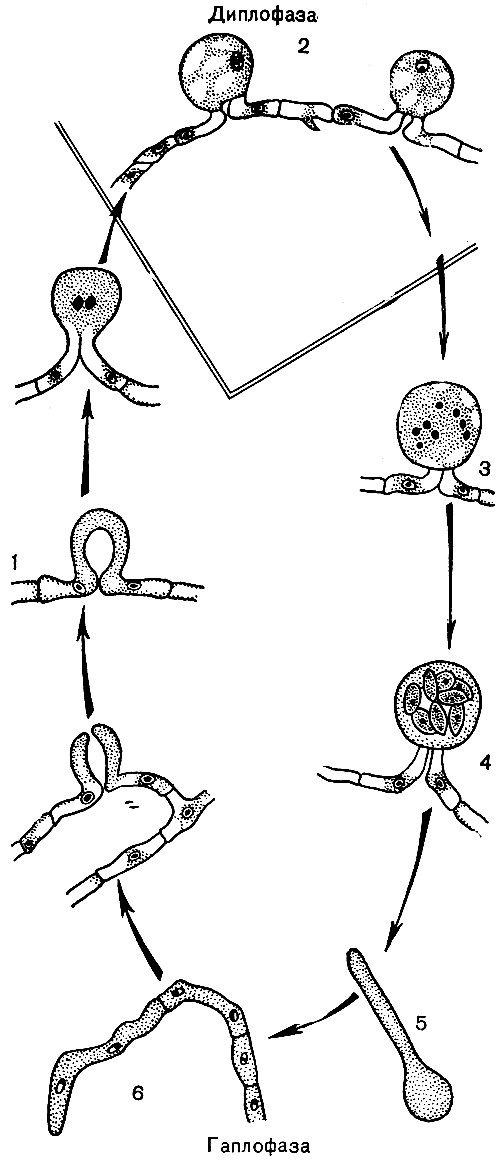
Рис. 52. Цикл развития гемиаскомицетов на примере эремаскуса фертильного (Eremascus fertilis): 1 - слияние гаментангие; 2 - зигота; 3 - молодая сумка с восемью гаплоидными ядрами; 4 - зрелая сумка с восемью аскоспорами; 5 - прорастающая аскоспора; 6 - гаплоидный мицелий
Для высших аскомицетов (подклассы Euascomycetidae и Loculoascomycetidae) характерны дифференциация и усложнение строения гаметангиев. Образуются одноклеточный антеридий и аскогон, обычно с трихогиной. При оплодотворении содержимое антеридия по трихогине переходит в аскогон. После плазмогамии гаплоидные ядра разного пола не сливаются сразу, а объединяются попарно, образуя дикарионы. Из аскогона вырастают аскогенные гифы, в которых ядра дикариона синхронно делятся. Аскогенные гифы ветвятся и разделяются на двухъядерные клетки. На концах аскогенных гиф развиваются сумки (см. рис. 50). Конечная клетка аскогенной гифы загибается крючком, ядра дикариона располагаются в месте перегиба и одновременно делятся. Пара ядер разного пола остается в месте перегиба крючка, одно ядро переходит в его кончик, а другое - в основание. Затем образуются две перегородки, отделяющие одноядерные конечную и базальную клетки крючка. В результате слияния этих клеток восстанавливается дикарион и может происходить повторное образование крючка. Средняя двухъядерная клетка крючка развивается в сумку. Она увеличивается в размерах, ядра дикариона сливаются. Образовавшееся диплоидное ядро делится редукционно, за мейозом следует еще одно, митотическое деление, и вокруг восьми гаплоидных ядер формируются аскоспоры.
По способу образования аскоспоры аскомицетов отличаются от спорангиоспор зародышевого спорангия зигомицетов. Первые образуются по способу так называемого "свободного образования клеток" - часть цитоплазмы сумки обособляется вокруг ядер и одевается оболочкой; при образовании спорангиоспор происходит раскалывание цитоплазмы спорангия. Аскоспоры в сумке окружены не использованной на их формирование цитоплазмой - эпиплазмой. К моменту созревания аскоспор в эпиплазме происходит превращение гликогена в сахар, тургорное давление в сумке возрастает (у некоторых видов - до 10-13 атм, по Ц. Ингольд, 1959) и аскоспоры с силой выбрасываются на расстояние от десятых долей миллиметра до нескольких десятков сантиметров.
Образование аскогенных гиф увеличивает число сумок, а следовательно, и аскоспор, развивающихся из одного аскогона. Образование сумок по способу крючка с восстановлением дикариона в его базальной клетке и повторным образованием крючков обеспечивает расположение сумок пучком или слоем, что очень важно при активном освобождении аскоспор.
У некоторых аскомицетов (например, из порядка эуроциевых) сумки образуются на аскогенных гифах иными способами - цепочками по ходу аскогенных гиф, из их конечных клеток или боковых выростов.
В цикле развития высших аскомицетов чередуются, таким образом, три стадии: длительная - гаплоидная, в течение которой происходит бесполое размножение, непродолжительная - дикарионтическая (аскогенные гифы) и очень короткая - диплоидная (молодая сумка с диплоидным ядром).
Для многих аскомицетов характерна морфологическая редукция полового процесса. У некоторых представителей этого класса антеридии отсутствуют или не функционируют. В этом случае их функции могут выполнять конидии, вегетативные гифы, а часто мелкие специализированные клетки, называемые спермациями. Спермации нередко образуются на другой особи, на значительном расстоянии от аскогона и переносятся на трихогину токами воздуха, дождем, насекомыми. Трихогина некоторых аскомицетов хемотропична и подрастает к спермациям или конидиям совместимого типа (Ascobolus stercorarius, Podospora anserina, виды Neurospora). Сперматизация обнаружена в разных группах аскомицетов, например у некоторых гелоциевых, видов из родов Ascobolus и Gelasinospora, в порядке лабульбениевых и у других.
В том случае, когда отсутствуют оба гаметангия, образование дикариона происходит в результате слияния клеток обычных вегетативных гиф одного или двух совместимых мицелиев - соматогамии (например, у Sclerotinia sclerotiorum).
Среди аскомицетов есть гомоталличные и гетероталличные виды, причем гетероталлизм здесь всегда биполярный (двухаллельного типа).
В сумке аскомицетов обычно образуется 8 аскоспор. Однако наблюдаются многочисленные отклонения от типа: например, происходит образование только четырех гаплоидных ядер и соответственно четырех аскоспор (Endomyces magnusii) или часть образовавшихся ядер дегенерирует и число аскоспор за счет этого сокращается (Verpa bohemica). Образование большего числа аскоспор происходит или при большем числе делений ядра (например, у Podospora - семь делений ядра и соответственно 128 аскоспор в сумке), или в результате почкования аскоспор в сумке (например, у тафриновых), или их распада на клетки (у некоторых гипокрейных).
Форма аскоспор очень разнообразна - от шаровидных или эллипсоидальных до нитевидных (Bhytisma, Claviceps). Аскоспоры могут быть одноклеточными или иметь поперечные перегородки, реже - поперечные и продольные перегородки (муральные споры).
У некоторых аскомицетов аскоспоры имеют разнообразные придатки, играющие роль в их распространении, например слизистые придатки аскоспор копрофильных грибов из рода Podospora или аскоспор сапрофитных морских аскомицетов Ceriosporiopsis lialima, Bemispora maritima и др.
По строению оболочки и функциям сумки аскомицетов делят на две большие группы - прототуникатные и эутуникатные. Прототуникатные сумки имеют тонкую недифференцированную оболочку, которая разрушается или растворяется, освобождая аскоспоры пассивно. Такая сумка служит только местом формирования спор, но не участвует активно в их распространении. Эутуникатные сумки характеризуются более плотными оболочками, часто со специальными приспособлениями для вскрывания сумки. Они активно участвуют в распространении аскоспор (активное выбрасывание спор из сумки). По строению оболочки эутуникатные сумки могут быть двух типов: унитуникатные и битуникатные. У унитуникатных сумок оболочка относительно тонкая и выглядит однослойной, на вершине сумок имеется обычно апикальный аппарат различного строения, служащий для их вскрывания. Битуникатные сумки имеют ясновыраженную двухслойную оболочку, состоящую из жесткого наружного и эластичного внутреннего слоев. При созревании аскоспор наружный слой оболочки разрушается, начиная с вершины; внутренний слой под действием повышенного тургорного давления растягивается, и происходит активное выбрасывание аскоспор.
Форма сумок округлая или овальная (у гемиаскомицетов и аскомицетов с замкнутыми плодовыми телами) или цилиндрическая (у аскомицетов с активным выбрасыванием аскоспор).
У низших аскомицетов сумки образуются непосредственно на мицелии, а у высших - в специальных вместилищах - плодовых телах (рис. 69). Различают следующие типы плодовых тел: клейстотеций (полностью замкнутое плодовое тело), перитеций (полузамкнутое, обычно кувшиновидное плодовое тело с отверстием на вершине) и апотеций (открытое, обычно чашевидное плодовое тело, на верхней стороне которого расположен слой сумок и парафиз). Клейстотеции, перитеции и апотеции - настоящие плодовые тела, развитие которых происходит по аскогимениальному типу - образование их оболочки (перидия) происходит одновременно с развитием аскогенных гиф и сумок.
Настоящие плодовые тела могут развиваться как непосредственно на мицелии, так и на стромах - плотных сплетениях гиф различной формы, размера и консистенции. Однако они всегда имеют собственный перидий, заметный хотя бы на ранних стадиях развития плодового тела.
Кроме настоящих плодовых тел, аскомицеты образуют аскостромы, развитие которых происходит по асколокулярному типу. Сначала закладывается строма из переплетающихся гиф. В ней образуются аскогоны и происходит половой процесс. Аскогенные гифы и образующиеся на них сумки раздвигают или разрушают плектенхиму стромы, освобождая в ней полость - локулу. Каждая локула содержит одну или несколько сумок. Ткань стромы над локулой разрушается и образуется отверстие, через которое освобождаются аскоспоры. По внешнему виду аскостромы часто похожи на настоящие перитеции, но отличаются от них отсутствием собственного перидия - их оболочкой служит плектенхима стромы.
На основании отсутствия или наличия плодовых тел и способов их образования класс аскомицеты делят на три подкласса:
Подкласс Голосумчатые, или Гемиаскомицеты (Hemiascomycetidae). Плодовые тела отсутствуют. Сумки образуются непосредственно на мицелии, прототуникатные.
Подкласс Эуаскомицеты (Euascomycetidae). Сумки образуются в клейстотециях, перитециях или апотециях, прототуникатные или унитуникатные.
Подкласс Локулоаскомицеты (Loculoascomycetidae). Сумки образуются в аскостромах, битуникатные.
Аскомицеты широко распространены в природе во всех географических областях, на всех доступных для грибов субстратах. Они обитают как сапрофиты в почве, в лесной подстилке, на разнообразных растительных субстратах (древесина, отмершие растения и т. п.). Некоторые группы аскомицетов заняли своеобразные экологические ниши, недоступные для других грибов. Например, кератинофильные грибы, развивающиеся на различных субстратах животного происхождения, содержащих кератин. Некоторые аскомицеты обитают в морях или пресных водоемах на погруженной в воду древесине. Сапрофитные аскомицеты активно участвуют в минерализации органических веществ в природе, особенно в разложении растительных остатков, содержащих целлюлозу. Многие сапрофитные аскомицеты вызывают плесневение и порчу разнообразных материалов и изделий, а также пищевых продуктов.
Многочисленные аскомицеты паразитируют на различных организмах - на растениях (грибах, водорослях, лишайниках и высших), а также на животных и человеке, нередко вызывают серьезные заболевания.
Многие представители этого класса имеют большое экономическое значение как продуценты антибиотиков, алкалоидов, ростовых веществ (гиббереллинов), витаминов (рибофлавина и др.), ферментов, кормового белка, а также как возбудители спиртового брожения. Наконец, многие аскомицеты широко используются сейчас в качестве объектов генетических и биохимических исследований.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'