Происхождение, родственные связи и эволюция водорослей (Ю. Е. Петров)
Вопрос о происхождении и эволюции водорослей очень сложен из-за разнообразия этих растений, особенно их субмикроскопического строения и биохимических признаков. Решение этой проблемы затруднено, кроме того, плохой сохранностью большинства водорослей в ископаемом состоянии и отсутствием связующих звеньев между современными отделами растений в виде организмов промежуточного строения.
Проще всего решается вопрос о происхождении прокариотических (доядерных) водорослей - сине-зеленых, имеющих много общих признаков с фотосинтезирующими бактериями. Вероятнее всего, сине-зеленые водоросли произошли от организмов, близких к пурпурным бактериям и содержащих хлорофилл а.
Относительно происхождения эукариотических (ядерных) водорослей в настоящее время нет единой точки зрения. Существуют две группы теорий, исходящих или из симбиотического, или из несимбиотического их происхождения.
По теории симбиогенеза хлоропласты и митохондрии клеток эукариотических организмов некогда были самостоятельными организмами: хлоропласты - прокариотическими водорослями, митохондрии - аэробными бактериями. Как предполагают, существовало несколько групп прокариотических водорослей, различающихся набором пигментов. В результате захвата амебоидными гетеротрофными эукариотическими организмами аэробных бактерий, и прокариотических водорослей возникли предки современных групп эукариотических водорослей. Некоторые исследователи приписывают симбиотическое происхождение хромосомам и жгутикам.
Теория симбиотического происхождения эукариотических водорослей и др. организмов получила особенно широкое распространение за последнее время. Она основана на давно известных фактах самовоспроизведения упомянутых органоидов клеток и на новых данных об автономности хлоропластов и митохондрий и их биохимическом сходстве с прокариотами. Это сходство выражается в следующем:
1. Хлоропласты и митохондрии, подобно прокариотам, содержат ДНК и РНК; в них находятся рибосомы того же типа, что и у прокарио-тов.
2. Антибиотики, подавляющие рост бактерий, тормозят образование и размножение хлоропластов и митохондрий эукариотических орга-низмов, но не действуют на рост самих клеток и тканей.
3. Механизмы фотосинтеза у сине-зеленых и эукариотических водорослей в значительной мере сходны.
Однако наряду с фактами, свидетельствующими в пользу симбиогенеза эукариотических организмов, имеются данные, говорящие об обратном. Например, система переноса электронов в хлоропластах и набор ферментов, необходимых для образования пигментов, участвующих в фотосинтезе, регулируются генами ядра. Синтез ряда ферментов митохондрий зависит от ядра и цитоплазмы. Если учесть эти и другие факты, хлоропласты и митохондрии оказываются не "автономными", а "полуавтономными". Что касается самовоспроизведения, то оно обнаружено даже у поверхностного (кортикального) слоя инфузорий, признаки которого наследуются в ряду поколений независимо от ядра и цитоплазмы. Это было установлено путем пересадок ядра и цитоплазмы других особей, обладающих кортикальным слоем с иными признаками. Конечно, никак нельзя представить, что кортикальный слой инфузорий некогда был самостоятельным организмом. На водорослях такие эксперименты не проводились.
Автономность хлоропластов и митохондрий в равной мере может свидетельствовать как об их некогда самостоятельном существовании, так и о далеко зашедшем процессе специализации и автономности органоидов клетки. Точно так же и данные о неполной автономности хлоропластов и митохондрий можно с равным успехом принять за доказательство их несимбиотического происхождения или рассматривать как пример утраты симбионтами полной автономности в ходе эволюции. Нам представляется, что частичная или полная автономность органоидов клетки от ядра является необходимым условием нормальной жизнедеятельности эукариотической клетки и своевременного реагирования ее органоидов на изменения окружающей среды. Эукариотические клетки в среднем в 103-104 раз крупнее прокариотических клеток. При полной зависимости органоидов от ядра малейшее нарушение его нормального функционирования, хотя бы на короткий отрезок времени, например при митозе, означало бы нарушение обмена веществ всей клетки.
Не в пользу симбиотического происхождения клеток эукариотических водорослей свидетельствует также отсутствие достаточного сходства в строении хлоропластов и клеток сине-зеленых водорослей - единственных современных прокариотических водорослей. Сейчас существует целый ряд бесцветных одноклеточных организмов, в которых живут в качестве симбионтов явные сине-зеленые водоросли, выполняющие функции хлоропластов и называемые цианеллами. Цианеллы отличаются от свободноживущих сине-зеленых водорослей. Интересно, например, что у симбиотического глаукоцистиса (Glaucocystis nostochinearnm), изученного посредством электронного микроскопа, цианеллы, в отличие от свободноживущих сине-зеленых водорослей, не имеют оболочки, т. е. симбиоз зашел здесь очень далеко. Тем не менее даже в таком "упростившемся" виде цианеллы все же остаются сине-зелеными водорослями и по своей организации и поведению в клетке хозяина решительно не сходны с хлоропластами (подробнее см. раздел "Сожительство водорослей с другими организмами").
Таким образом, существующие ныне данные не позволяют принять теорию симбиогенеза в качестве основы для пересмотра путей возникновения и эволюции эукариотических водорослей.
Если исходить из несимбиотического происхождения эукариотических водорослей, то приходится допустить, что они возникли от предка, общего с сине-зелеными водорослями, имеющего хлорофилл а и фотосинтез с выделением кислорода (рис. 272). Единственным возражением против этого может быть разница в составе клеточной стенки: у сине-зеленых водорослей, так же как и у бактерий, в клеточной стенке имеется муреин. В целом по составу и строению клеточной стенки, а также по реакциям, благодаря которым идет синтез ее веществ, прокариоты существенно отличаются и от животных, и от остальных растений. В случае принятия такого возражения пришлось бы выводить эукариотические водоросли от других организмов. Это значило бы признать, что фотосинтез с участием хлорофилла а и выделением кислорода возникал в ходе эволюции жизни на Земле два раза. Однако это, учитывая множество реакций, осуществляемых в процессе фотосинтеза с участием многих ферментов, представляется менее вероятным, чем смена в ходе эволюции веществ клеточной стенки. У эукариотических водорослей, очевидно, не сразу появилась твердая жесткая клеточная стенка из целлюлозы или других веществ. Наиболее примитивной у эукариотических водорослей, очевидно, следует считать амебоидную форму строения, а клеточная стенка всех современных прокариот имеет жесткую основу. Следовательно, современные фотосинтезирующие прокариоты, т. е. сине-зеленые водоросли,- это боковая, тупиковая ветвь растительной эволюции. Эукариотические водоросли имели с ней лишь общего прокариотического предка, лишенного твердой оболочки.
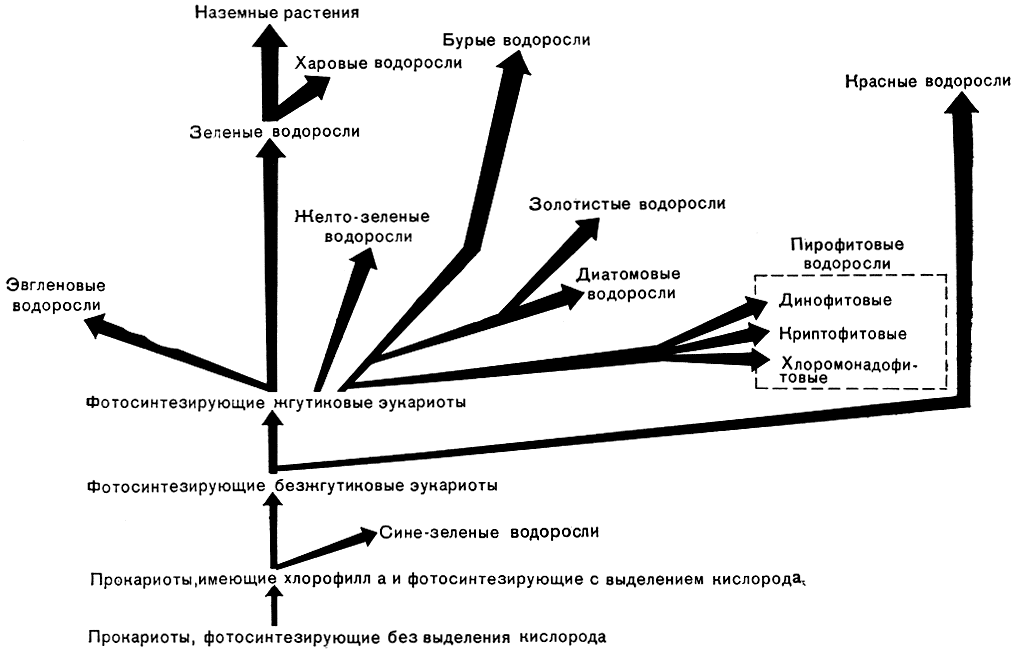
Рис. 272. Схема происхождения и эволюционного развития водорослей
Первым шагом на пути возникновения эукариотических водорослей было формирование ядра и хлоропластов. На этом этапе эволюции появились красные водоросли, для которых характерно отсутствие жгутиковых стадий, примитивность строения хлоропластов и появление второй разновидности хлорофилла - хлорофилла d. Несмотря на сложность цикла развития и анатомического строения, достигнутую некоторыми представителями, красные водоросли оказались боковой и слепой ветвью эволюции.
Важнейшим моментом в становлении органического мира на Земле явилось развитие у одноклеточных эукариотов двигательного аппарата - жгутиков, построенных весьма своеобразно: внутри их по периферии располагаются 9 пар фибрилл и 2 фибриллы находятся в центре. С их появлением зародилась центральная прогрессивная группа - фотосинтезирующие эукариотические жгутиковые. Начиная с этого этапа эволюция эукариотов пошла в нескольких направлениях: у одних организмов в дополнение к хлорофиллу а появились хлорофилл b, с или е и ряд новых дополнительных пигментов, а другие организмы утратили фотосинтезирующие пигменты и полностью перешли к гетеротрофному питанию. Одновременно происходила эволюция, сопровождающаяся видоизменением первоначального строения жгутикового аппарата. Например, у диатомовых исчезла центральная пара фибрилл, у некоторых золотистых водорослей появилась гаптонема (уплощенный "жгутик") с 5-8 одиночными фибриллами по периферии и тремя мембранами в оболочке. Правда, некоторые исследователи допускают, что эукариотические водоросли могли возникнуть от гетеротрофных безжгутиковых эукариотических организмов. Но это было бы возможно, если допустить, что фотосинтез с выделением кислорода и жгутики, имеющие одно и то же исходное строение у всех водорослей и гетеротрофных организмов, появлялись дважды. Существуют также предположения, что сине-зеленые водоросли непосредственно дали начало красным водорослям, а водоросли, обладающие жгутиками, произошли от бесцветных жгутиковых. Однако это означало бы, что дважды в ходе эволюции формировалось ядро, имеющее одно и то же строение и делящееся митотически, и дважды создавались одинаковые фотосинтезирующие системы с хлорофиллом а и хлоропластами. Все это крайне маловероятно.
Согласно изложенным представлениям о путях возникновения эукариотических водорослей, гетеротрофные жгутиковые приходится выводить из фотосинтезирующих самостоятельно в нескольких ветвях. Проще допустить неоднократную утрату хлоропластов, которая, к то-му же, действительно наблюдается у представителей многих групп одноклеточных водорослей, чем предположить симбиотическое происхождение эукариот или многократное возникновение фотосинтеза и единообразно устроенных у всех эукариот жгутиков и ядер. Утрата пластид некоторыми жгутиковыми тем более допустима, что фототрофные организмы, как прокариоты, так и эукариоты, никогда не теряли полностью способности к гетеротрофному питанию, унаследованному от первичных гетеротрофов, появившихся при зарождении жизни на Земле. Вместе с тем всем организмам, живущим на Земле, свойственно биохимическое единство независимо от способа питания (автотрофного или гетеротрофного). Дж. Бернал, например, писал: "...при рассмотрении биохимии в целом, включая процессы, протекающие у всех видов животных и растений, а также бактерий и вирусов, обнаруживалось необычайное единство и экономичность. Все снова и снова мы встречаемся с одними и теми же химическими реакциями и структурами - вплоть до деталей атомной структуры. И даже там, где наблюдаются вариации, это вариации на одну и ту же тему. Так, например, порфирины используются в дыхательных ферментах, при фотосинтезе и при переносе кислорода у высших животных".
Первичные фотосинтезирующие жгутиковые эволюционировали по двум главным направлениям. У одних жгутиковых преобладающее значение получили бурые пигменты, и у ряда представителей появился второй хлорофилл - с или е, а в числе запасных питательных веществ у многих представителей стали образовываться разные модификации специфического полисахарида ламинарина. Второе направление эволюции жгутиковых характеризуется преобладанием зеленой окраски и наличием хлорофилла b наряду с хлорофиллом а. Промежуточное положение между этими двумя основными направлениями заняли жгутиковые, которые, эволюционируя, привели к появлению разнородной группы желто-зеленых водорослей, имеющих зеленую окраску. Хлоропласты желто-зеленых водорослей не содержат хлорофилла b и имеют такое же строение, как у большинства водорослей с бурыми пигментами. В качестве запасного вещества может образовываться хризоламинарин; крахмал и парамилон отсутствуют. В пределах каждого из этих направлений эволюция пошла по нескольким линиям, нередко параллельно, сопровождаясь возникновением сходных форм строения тела.
Жгутиковые с преобладанием бурых пигментов эволюционировали несколькими путями, которые привели к возникновению водорослей: золотистых, диатомовых, бурых и пирофитовых. Последние фактически представляют два самостоятельных отдела, из которых один, объединяющий динофитовые водоросли, выделяется строением ядерного аппарата не только среди водорослей с бурыми пигментами, но и среди всех эукариотических организмов. Из числа водорослей, окрашенных в бурый цвет, наибольшего расцвета достигли диатомовые и бурые. Диатомовые заняли доминирующее положение среди микроскопических водорослей морей и континентальных водоемов. Бурые водоросли оказались наиболее приспособленными к жизни в прибрежной зоне моря. У них возник ряд приспособлений, позднее появившихся на иной основе у наземных растений. Здесь имеется в виду наличие у бурых водорослей ситовидных трубок, подобных по субмикроскопическому строению проводящим элементам флоэмы выс-ших растений, а также развитие спорофита на гаметофите или гаметофита на спорофите и размножение некоторых видов посредством сформированных проростков спорофитов. Бурые водоросли захватили в море наиболее благоприятные для роста донных водорослей места, где существует твердый грунт и почти постоянное движение воды. Среди морских донных водорослей бурые водоросли заняли такое же положение, как покрытосеменные растения на суше. В планктоне морей значительную долю растительных организмов, кроме диатомовых, составляют еще только кокколитофориды из золотистых водорослей и динофитовые водоросли.
Эволюция жгутиковых с зелеными пигментами шла в основном в одном направлении, охватывающем отдел зеленых водорослей. От него очень рано отделились эвгленовые водоросли, которые не эволюционировали дальше одноклеточного состояния. От многоклеточных зеленых водорослей в качестве боковой ветви эволюции отделились харовые водоросли. У них наиболее сложно устроенные органы полового размножения среди водорослей с зелеными пигментами, и тем не менее это тупиковая группа, представленная сейчас ограниченным числом родов и видов довольно однотипного строения. В прошлые геологические эпохи пышного развития достигали из зеленых водорослей дазикладовые и сифоновые, последние и сейчас играют существенную роль в жизни тропических морей. Эволюция зеленых водорослей в водной среде не привела к появлению сложно устроенных и крупных слоевищ тканевого строения, сравнимых с бурыми и красными водорослями. Продолжением эволюции зеленых водорослей явилось возникновение фотосинтезирующих высших наземных растений.
Происхождение наземных растений иногда пытаются связывать с бурыми водорослями из-за существования у них многогнездных спорангиев и гаметангиев, которые ошибочно принимают за многоклеточные органы размножения. Вместилища бурых водорослей делятся на камеры в последний момент при формировании в них зооидов, чего не наблюдается у наземных растений. В то же время бурые водоросли сильно отличаются от наземных растений набором пигментов, запасными питательными веществами, химическим составом оболочки.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'