Порядок церамиевые (Ceramiales)
По объему церамиевые намного превосходят все другие порядки в классе флоридеевых. Достаточно сказать, что они объединяют более 250 родов и около 1500 видов. Но, несмотря на столь большой объем, порядок - хорошо очерченная группа, которая делится всего на 4 четко ограниченных семейства. Единообразие порядка проявляется прежде всего в структуре и в развитии органов размножения. Всем церамиевым свойствен прокарп. Ауксиллярная клетка отчленяется непосредственно от несущей клетки четырехклеточной карпогонной ветви. Формируется ауксиллярная клетка только после оплодотворения. Цикл развития, как правило, трехфазный со сменой гаметофита, карпоспорофита и тетраспорофита; при этом первая и последняя формы развития имеют одинаковое строение. В то время как репродуктивная система церамиевых построена по единому плану, вегетативная организация их отличается разнообразием. На этом и основана классификация порядка. Все церамиевые построены по одноосевому типу.
Из четырех семейств на самой низкой ступени развития стоит семейство церамиевых. Остальные три, по-видимому, представляют параллельные линии эволюции, и сейчас трудно сказать, какое из них стоит на более высокой ступени организации.
Распространены церамиевые по всему Мировому океану, но особенно богато представлены в тропиках.
Семейство церамиевые (Ceramiaceae)
Обширное семейство церамиевых включает в себя четвертую часть всех родов и видов порядка. В большинстве своем водоросли семейства - мелкие, нежные организмы нитчатой или цилиндрической формы, лишенные коры или покрытые ею. В отличие от других у пред-ставителей этого семейства структура никогда не бывает полисифонной. Кроме того, церамиевые имеют голые гонимобласты. Если обертка и формируется, то она состоит из коротких веточек, развивающихся отдельно от гонимобласта. У остальных водорослей порядка обертка плотного паренхимного строения.
Простейшую организацию мы находим у родов круания и антитамнион.
Род круания (Crouania), включающий 10 видов, наиболее богато представлен у берегов Австралии, хотя типовой вид рода широко распространен у Атлантического побережья Европы и в Средиземном море. Нежные слизистые растения состоят из однорядной оси, покрытой мутовками коротких, многократно разветвленных изогнутых веточек. Каждая мутовка состоит из 3-4 веточек. Развитие четвертой ветви может подавляться или из нее вырастает ветвь неограниченного роста (возникновение основных ветвей вместо укороченных - примитивная черта). Формирование ветвей в отдельной мутовке не отличается от такового у других церамиевых. Вторая всегда закладывается напротив первой, третья - между ними, четвертая - напротив третьей. У всех церамиевых первые ветви мутовок вдоль оси слоевища располагаются по спирали. Место их возникновения и, следовательно, интервалы, через которые они следуют друг за другом, строго зафиксированы для каждого рода. У круании они закладываются таким образом, что ветви соседних мутовок смещены на оси по отношению друг к другу на 45° (полный оборот спирали составляет 360°). Боковые веточки резко отличаются от основных ветвей толщиной и ветвлением. Первые ветвятся дихотомически и политомически, вторые - моноподиально (рис. 182, 1).
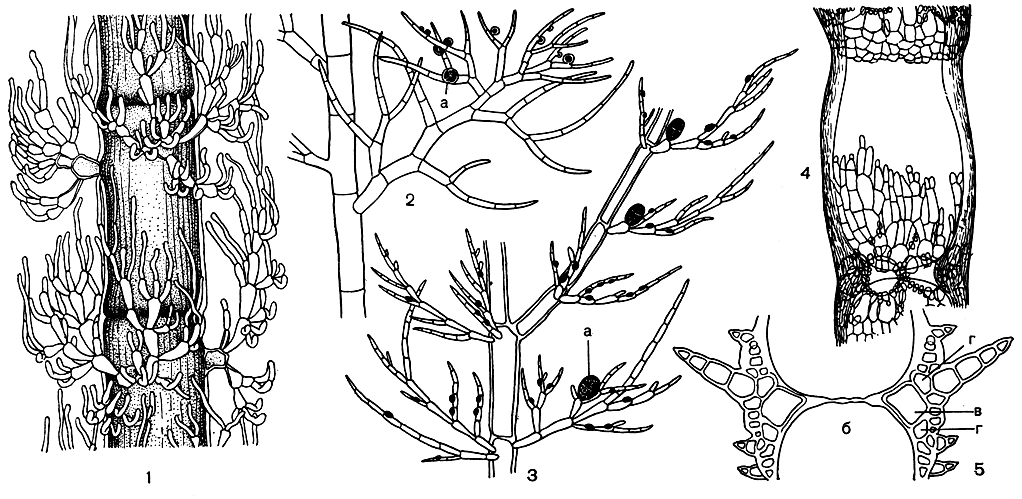
Рис. 182. Строение церамиевых: 1 - часть веточки Crouania; 2 - веточка Callithamnion; 3 - веточка Antithamnion; 4 - часть слоевища Ceramium; 5 - продольный срез через узел Ceramium. а - тетраспоры, б - центральная клетка, в - перицентральная клетка, г - вторичные коровые клетки
По особенностям строения женской половой системы и карпоспорофита круания стоит на низкой ступени развития. В зрелом слоевище одна из перицентральных клеток формируется как несущая и вместо обычной дает начало карпогонной ветви. Последняя полностью замещает вегетативную, никаких стерильных клеток рядом с ней не остается (рис. 183, 1). Сперматангии образуются на вершине обычных веточек ограниченного роста. Тетраэдрически разделенные тетраспорангии сидят на нижних клетках боковых ветвей и разбросаны по всему слоевищу.
Род каллитамнион (Callithamnion) близок к круании, но уже гораздо более специализирован. Известно более 50 видов каллитамниона, распространенных в различных морях. Несколько видов обитает в Черном море, среди них наиболее часто встречается каллитамнион щитковидный. У каллитамниона на каждом сегменте образуется по одной ветви, никаких следов мутовчатого ветвления в вегетативных ветвях обнаружить не удается (рис. 182, 2). Оно проявляется только в фертильных веточках. Объяснить характер ветвления каллитамниона и его происхождение в настоящее время весьма затруднительно. Некоторые виды имеют кору, которая состоит из ризоидных нитей, спускающихся по стенкам основных ветвей слоевища.
Карпогонные ветви образуются попарно из двух перицентральных клеток, возникающих вокруг основной оси. Обе фертильные перицентральные клетки представляют собой остатки ветвей ограниченного роста и вместе с вегетативной ветвью являются единственным проявлением мутовчатого ветвления у каллитамниона.
Род антитамнион (Antithamnion) также характеризуется простой организацией. Как и круания, эти водоросли построены однорядными лишенными коры и свободно разветвленными нитями. Ветви ограниченного роста отходят по две супротивно или по три-четыре му-товками. Вдоль основной оси все ветви ограниченного роста располагаются строго друг под другом, а не смещены, как у круании. По внешнему виду они почти не отличаются от основных ветвей. И те и другие ветвятся моноподиально; в боковых ветвях также прослеживаются основная ось и боковые ветви. Характерным признаком многих видов служит наличие железистых клеток на коротких веточках (рис. 182, 3).
Как у круании, карпогонная ветвь у антитамниона берет начало от перицентральной клетки, но здесь она не полностью замещает вегетативную, а занимает боковое положение на обычной веточке ограниченного роста. Зрелый гонимобласт окружен многочисленными соседними вегетативными веточками (рис. 183, 2, 3). Гаметофиты двудомные. Тетраспорофиты встречаются в природе чаще, чем гаметофиты. На боковых веточках они несут крестообразно поделенные тетраспорангии.
Род церамиум (Ceramium) относится к той же группе родов, что и антитамнион. Около 60 видов церамиума распространено от тропиков до арктических морей. Наибольшего видового разнообразия род достигает в теплых морях. Церамиум интересен как водоросль, имеющая кору. У некоторых видов кора покрывает сплошь все слоевище; другие же имеют развитый коровой слой лишь на отдельных участках, которые повторяются через определенные интервалы, придавая растению членистый вид (рис. 182, 4). Покрытые корой окрашенные "узлы" чередуются с голыми прозрачными "междоузлиями". Эту членистость часто можно заметить невооруженным глазом и по ней легко отличить церамиум от других багрянок. Характерной чертой в облике церамиума является также дихотомическое ветвление и вильчато-разделенные вершинки (табл. 25).

Таблица 25. Красные и бурые нитчатые водоросли: вверху - десмарестия зеленеющая (Desmarestia viridis); внизу - церамиум красйый (Ceramium rubrum) и другие мелкие багрянки на мидиях. Фото Ю. Астафьева
Как же построено слоевище церамиума? Центральная ось состоит из одного ряда крупных бесцветных клеток. Делясь косыми перегородками, они отчленяют вбок перицентральные, которые являются первичными коровыми клетками. Они располагаются кольцом из 6-10 довольно крупных клеток вокруг центральной оси. Из первичных клеток коры развиваются вверх и вниз по слоевищу мелкоклеточные нити, которые образуют внешний коровой слой. Клетки центральной оси при этом вытягиваются (рис. 182, 5). Если коровые нити, развивающиеся от двух соседних осевых клеток, смыкаются, образуется сплошной коровой слой, но чаще между ними остается свободный участок сегмента. Таким образом, коровые нити церамиума соответствуют веточкам ограниченного роста антитамниона, а первичные коровые клетки - нижним клеткам этих веточек. Сходство структур подтверждается и тем, что прокарп у церамиума возникает на одной из первичных коровых клеток. Церамиум - один из высокоорганизованных родов семейства. По-видимому, он происходит от каких-то антитамниевых в результате увеличения числа веточек ограниченного роста в мутовке и более густого их расположения.
Гаметофиты церамиума в природе встречаются гораздо реже, чем тетраспорофиты, поэтому признаки половых растений меньше используются в систематике рода, чем признаки спорофитов. Сперматангии развиваются из поверхностных коровых клеток и, когда их много, как муфтой одевают "узлы" (рис. 183, 4). Возникновение прокарпов приурочено к вершинам длинных ветвей слоевища. Карпогонная ветвь располагается сбоку первичной коровой клетки. От нее же верхушечно отходит группа стерильных клеток - результат редукции обычной веточки ограниченного роста. Зрелые гонимобласты окружены короткими вегетативными веточками, расположенными розеткой и образующими обертку цистокарпа (рис. 183, 5). Тетраэдрически разделенные спорангии развиваются из коровых клеток и располагаются по-разному: одиночно или кольцеобразно на сочленениях, выступая из коры или обрастая ею (рис. 183, 6). У видов со сплошной корой тетраспорангии разбросаны по всему слоевищу. Все эти признаки важны для разграничения видов.
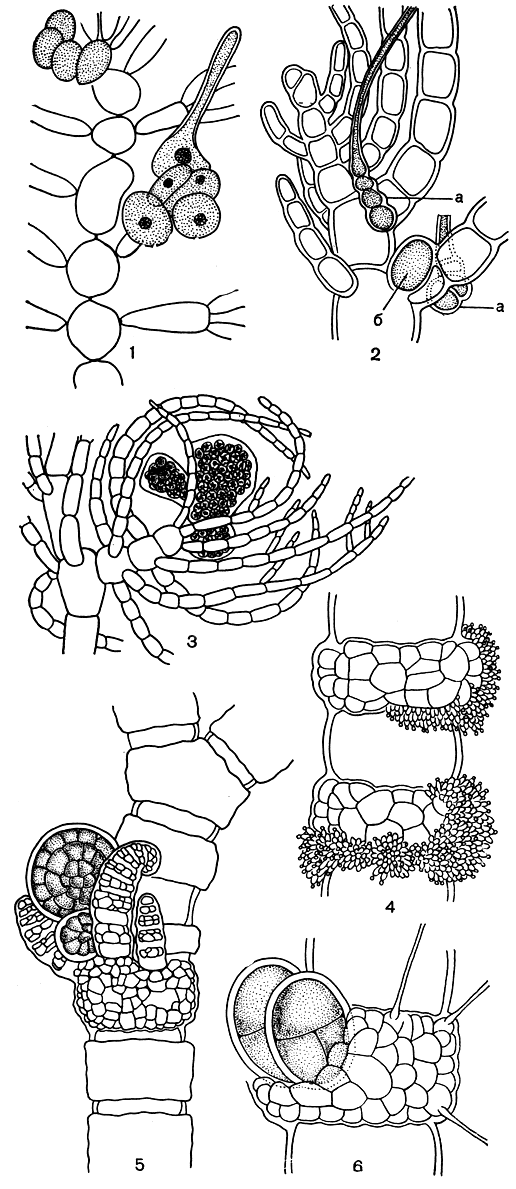
Рис. 183. Размножение церамиевых: 1 - прокарп Crouania; 2 - прокарп Antithamnion; 3 - зрелый цистокарп Antithamnion; 4 - сперматангии Ceramium; 5 - молодой цистокарп Ceramium; 6 - выступающие тетраспорангии Ceramium. а - карпогонная ветвь, б - ауксиллярная клетка
Семейство дазиевые (Dasyaceae)
Это небольшая группа, насчитывающая 12 родов, приуроченных к тропической и субтропической зонам. В моря умеренной зоны заходят лишь немногие виды. Среди наиболее обширных и широко распространенных родов следует назвать дазию (Dasya) и гетеросифонию (Heterosiphonia). Они же представляют интерес для понимания особенностей строения и размножения всего семейства. Главное отличие дазиевых от остальных водорослей порядка заключается в симподиальной организации. Напомним, что симподиальное ветвление характеризуется тем, что в месте отхождения ветви главная ось смещается в сторону, а ее место занимает вновь образовавшаяся ветвь, т. е. постоянной апикальной клетки здесь нет, а ее роль принимают на себя апикальные клетки последовательно возникающих ветвей (рис. 184, 1). Слоевище дазии имеет радиальную симметрию. Главные ветви покрыты густо разветвленными однорядными веточками ограниченного роста, что придает растениям опушенный вид (табл. 23, 4). Отходят эти веточки по одной от каждого сегмента и располагаются по спирали.
Апикальная клетка отчленяет от себя осевую клетку, последняя быстро растет, смещая апикальную клетку в сторону, и сама занимает ее место. Вот эти-то отошедшие в сторону апикальные клетки и дают начало однорядным боковым веточкам. По внешнему виду они напоминают трихобласты родомеловых, но по способу возникновения не имеют с ними ничего общего. Сегменты основных ветвей очень скоро становятся многорядными, так как отчленяют пять перицентральных клеток, или сифонов. Их образование оказывается совершенно иным, чем у родомеловых. На клетках центральной оси, в месте возникновения перицентральной клетки, образуется почковидный выступ, который позднее отделяется перегородкой. Первоначально перицентральные клетки короче центральных, у родомеловых они того же размера. Порядок, в котором происходит отчленение перицентральных клеток, также отличен от родомеловых. Из нижних частей перицентральных клеток вырастают мелкоклеточные разветвленные ризоидные нити, которые, смыкаясь, формируют многорядную плотную кору (рис. 184, 2). В образовании коры также принимают участие ризоиды, берущие начало от основания коротких боковых веточек. Между собой клетки коры соединяются вторичными порами. Строение слоевища дазиевых на ранних стадиях прорастания совсем иное, чем у родомеловых. Проростки дазиевых однорядные, нитчатые, свободно разветвленные, тогда как у родомеловых - многорядные и неразветвленные.
Прокарп у дазии закладывается близ вершины основных ветвей или на дополнительных боковых веточках. Фертильным обычно становится третий из последовательно возникающих перицентральных сегментов. Его можно отличить уже в самом начале - он крупнее остальных и более богат протоплазматическим содержимым. Эта клетка служит несущей и дает начало карпогонной ветви и двум стерильным клеткам. До оплодотворения обе стерильные клетки почти не отличаются от вегетативных и не подвергаются делению. После оплодотворения несущая клетка дает начало ауксиллярной, которая сливается с карпогоном и образует нити гонимобласта. В карпоспоры превращаются несколько верхних клеток этих нитей. Зрелый гонимобласт находится в полости, окруженной оберткой паренхимного строения. Так же как у делессериевых, формирование обертки начинается только после оплодотворения и происходит за счет разрастания соседних с фертильной перицентральных клеток. У родомеловых обертка закладывается еще до оплодотворения. Тетраэдрически-разделенные спорангии развиваются в специальных коротких веточках - стихидиях, располагаясь поперечными рядами среди их клеток (рис. 184, 3).
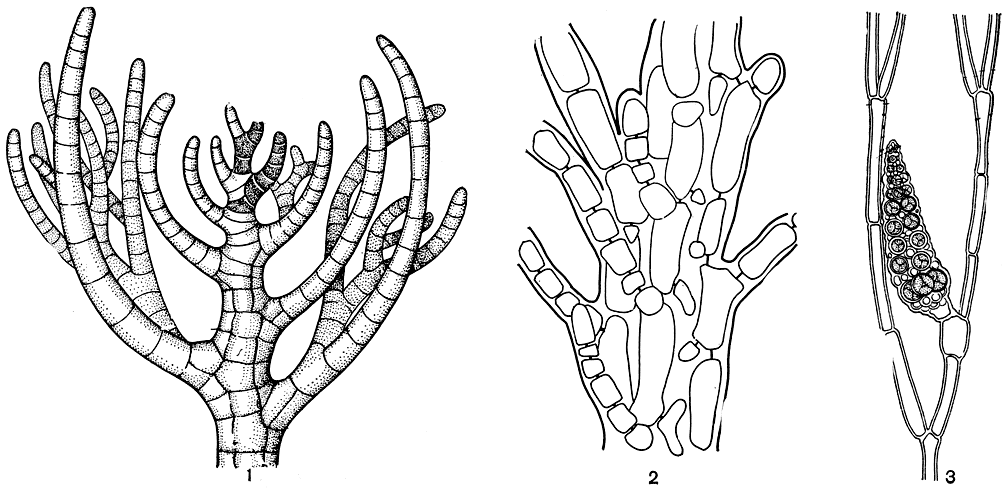
Рис. 184. Строение дазиевых: 1 - вершина веточки Heterosiphonia; 2 - образование коры у Dasya; 3 - стихидий Dasya с тетраспорангиями
Семейство делессериевые (Delesseriaceae)
Представители семейства делессериевых - одни из самых красивых водорослей, и не только среди багрянок, но, пожалуй, и среди всех водорослей-макрофитов. Растущие на глубине, они обычно окрашены в яркие красные тона. Нежные полупрозрачные пластины многих делессериевых пронизаны нервами и жилками и этим очень напоминают листья наземных растений. Сходство с листьями особенно велико у водорослей с изрезанным или зубчатым краем пластины. Когда по мере роста пластины разрушаются, остающиеся нервы можно принять за веточки, на которых сидят молодые листочки (табл. 22, 1, 3; 23, 2). Делессериевые красивы как в живом состоянии, только что взятые из моря, так и на бумаге, в расправленном и засушенном виде. Особенно красивы зрелые растения - с развитием тетраспор и цистокарпов пластины становятся бахромчатыми и покрываются темными пятнышками. Семейство включает большое число сублиторальных форм, которые распространены по всему Мировому океану, но наибольшего развития достигают в морях южного полушария.
Слоевище делессериевых построено по одноосевому типу. Ветви ограниченного роста, отходящие от центральной оси, развиваются в одной плоскости, что приводит к образованию пластинчатого слоевища, в отличие от других водорослей порядка. Рост осуществляется апикальной клеткой или краевой меристемой. Для многих характерны интеркалярные клеточные деления.
70 родов делессериевых объединяют в два подсемейства.
Подсемейство делессериевые (Delesserioideae)
Род делессерия (Delesseria) - типичный представитель подсемейства. Листовидное однослойное слоевище делессерии пронизано средним нервом и многочисленными боковыми жилками. Новые пластинчатые ветви возникают от среднего нерва, а не от края пластины, как это происходит у некоторых других родов.
Как же построено слоевище делессерии? Если посмотреть на молодую пластину с поверхности (рис. 185,1), то в ней можно хорошо различить апикальную клетку, которая отчленяет вниз сегменты центральной оси. От центральных сегментов берут начало четыре перицентральные клетки. Две из них, боковые, расположенные супротивно, развиваются в клеточные разветвленные нити, за счет которых строится пластина. Среди них можно различить ответвления первого, второго и более высоких порядков. Ветви первого порядка всегда достигают края пластины, тогда как ветви последующих порядков могут кончаться внутри слоевища. Как растут боковые нити, какое количество порядков они образуют, какие из них достигают края пластины - все это важные родовые признаки. Клетки центральной оси, раз образовавшись, уже не подвергаются поперечным клеточным делениям, а лишь вытягиваются в длину. Клетки боковых нитей растут так же, как и центральная, но, в отличие от нее, в них встречаются интеркалярные деления. В результате такого роста у делессерии четко прослеживается центральная ось, но расположение клеток в боковых нитях оказывается менее упорядоченным. Отсутствие интеркалярных делений в центральной оси - признак, отличающий все роды подсемейства. У многих из них интеркалярные деления отсутствуют и в боковых нитях. Поэтому представителей подсемейства всегда легко отличить по упорядоченному расположению клеток и отчетливой апикальной клетке. Две другие перицентральные клетки не получают такого развития. Они формируют кору над центральной осью. За счет этой разрастающейся коры и многочисленных ризоидных нитей, берущих начало от клеток коры, образуется утолщенный срединный нерв, пронизывающий слоевище делессерии.
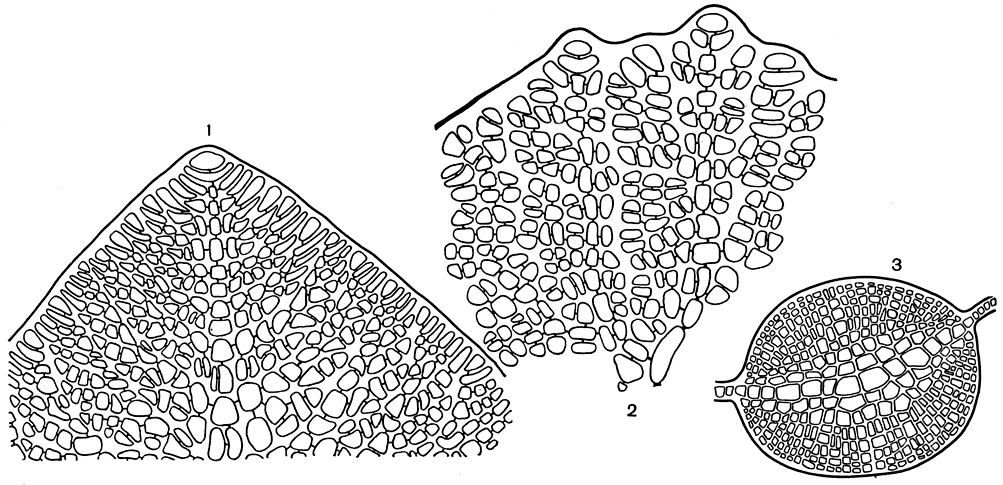
Рис. 185. Строение делессериевых: 1 - вершина слоевища Delesseria; 2 - вершина слоевища Phycodrys; 3 - поперечный срез через ребро Phycodrys
На зрелых растениях можно увидеть маленькие листочки - в них вдоль среднего нерва образуются прокарпы. Несущими клетками служат перицентральные клетки главной оси плодоносного листочка. Кроме карпогонной ветви, на несущей клетке образуются две группы стерильных клеток. После оплодотворения от нее же отделяется ауксиллярная клетка. Стерильные клетки делятся и наполняются густым содержимым, однако в определенный период оболочки их набухают и ослизняются, а сами клетки постепенно исчезают. В процессе развития гонимобласта из центральной, несущей и ауксиллярной клеток образуется клетка слияния, которая служит для питания развивающегося гонимобласта.
Стенки цистокарпов формируются в результате разрастания клеток коры. Большинство клеток гонимобласта превращается в карпоспоры. Образование сперматангиев и тетраспор также приурочено к особым мелким листочкам.
Подсемейство нитофилловые (Nitophylloideae)
Род фикодрис (Phycodrys) включает виды, внешне очень напоминающие делессерию. Так же как у делессерии, листовидное слоевище фикодриса пронизано средним нервом и боковыми жилками, но новые пластины вырастают по краю старой пластины в результате разрастания ее лопастей или роста боковых жилок. Основные различия между двумя родами и соответственно между двумя подсемействами проявляются в анатомическом строении и размножении. Слоевище фикодриса строится по общему для всех делессериевых принципу, но у него клетки центральной оси легко подвергаются интеркалярным клеточным делениям, в результате чего порядок в расположении клеток нарушается и уже в молодых пластинах проследить центральную ось и боковые нити, в отличие от подсемейства делессериевых, не удается (рис. 185, 2). За счет разрастания коровых клеток вдоль центральной оси слоевища у фикодриса образуется ребро, жилки приурочены к боковым ветвям первого порядка. Ризоиды здесь отсутствуют и участия в формировании ребра не принимают (рис. 185, 3).
У некоторых нитофилловых апикальный рост заменяется ростом за счет краевой меристемы. Для таких форм характерно вовсе беспорядочное расположение клеток и отсутствие апикальной клетки, как, например, у нитофиллума (Nitophyllum). Прокарпы в обоих подсемействах построены по единому плану, но у фикодриса они разбросаны по всей пластине. Точно так же сперматангии и тетраспорангии, собранные в сорусы, расположены на пластинчатой части, но не на ребрах и жилках.
Семейство родомеловые (Rhodomelaceae)
Это семейство, включающее более 100 родов,- самое крупное во всем отделе красных водорослей. Оно характеризуется огромным разнообразием вегетативных форм и в то же время поразительным однообразием в строении органов размножения. В существующей клас-сификации родомеловые занимают место наиболее высокоорганизованной группы отдела.
Род полисифония (Polysiphonia) - прекрасный пример структуры и размножения родомеловых. В тонконитчатом разветвленном слоевище полисифонии клетки располагаются правильными ярусами, или члениками. Каждый членик состоит из центральной клетки и расположенных вокруг нее нескольких перицентральных, одинаковой формы и длины (рис. 187,2). Такую структуру называют полисифонной. Число перицентральных клеток постоянно для каждого вида, но в пределах рода может заметно меняться.
Рост у полисифонии апикальный. Если образуется неветвящаяся ветвь, апикальная клетка делится поперечной перегородкой, отчленяя центральные сегменты. Образовавшиеся сегменты подвергаются продольным делениям, которые происходят в строгой последовательности - направо и налево от первой перицентральной клетки. В результате образуется полисифонный членик. Когда образуется разветвленная ветвь, апикальная клетка делится косо и образовавшийся клиновидный сегмент дает вырост, который является началом боковой ветви. Только после этого центральный сегмент делится продольно с образованием перицентральных клеток (рис. 186). Возникающая ветвь может быть ветвью неограниченного роста, и тогда она проходит то же развитие, что и главная ось, или короткой ветвью ограниченного роста, оставаясь при этом частично или полностью однорядной. Короткие однорядные веточки - трихобласты - располагаются на слоевище по спирали в строгой последовательности и через определенные интервалы, при этом в каждом членике образуется только один трихобласт. Обычно трихобласты вскоре после образования опадают, оставляя лишь маленькую базальную клетку (рис. 187,2). Длинные ветви неограниченного роста могут возникать в пазухе базальных клеток трихобластов или непосредственно от них. Слоевище многих видов полисифонии, как и других родов семейства, покрыто корой. Она развивается в результате деления перицентральных клеток или образуется возникающими от них же ризоидами.
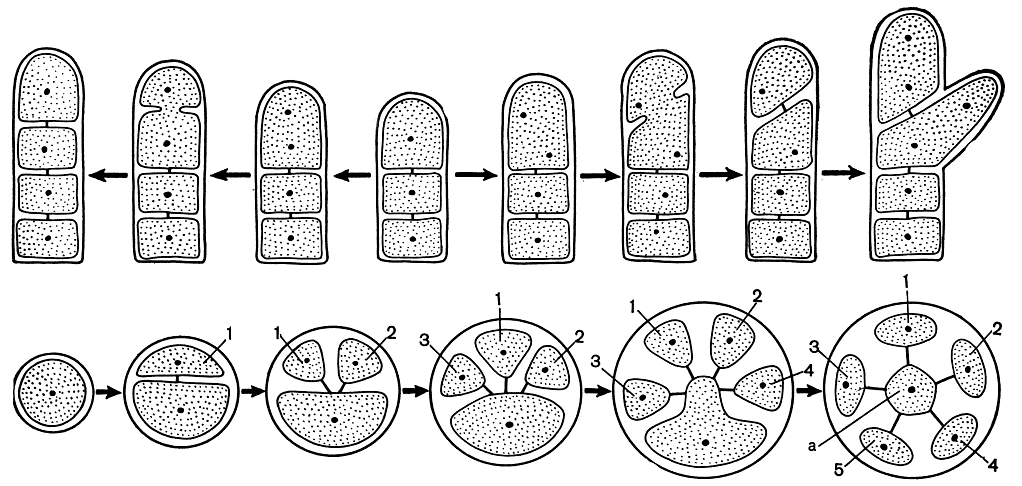
Рис. 186. Схема строения верхушки слоевища Polysiphonia: вверху - деление апикальной клетки при образовании разветвленных и неразветвленных сегментов; внизу - образование перицентральных клеток (поперечный срез), а - центральный сифон, 1-5 - перицентральные клетки
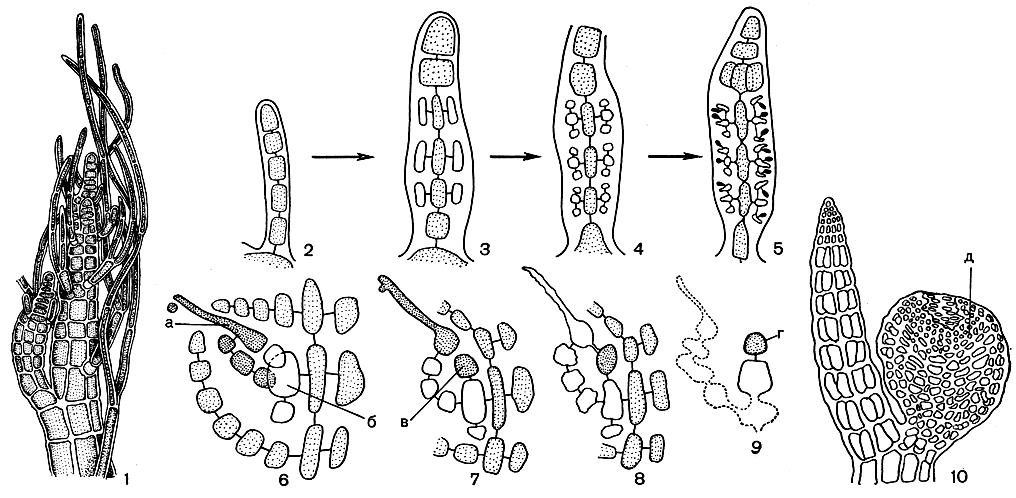
Рис. 187. Строение и размножение Polysiphonia: 1 - вершина веточки; 2-5 - схема развития сперматангиев на трихобласте; 6-10 - схема развития гонимобласта. а - карпогон, б - несущая клетка, в - ауксиллярная клетка, г - первая клетка гонимобласта, д - цистокарп
Органы полового размножения располагаются на плодоносных трихобластах. Прокарп всегда образуется на втором от основания сегменте трихобласта, который становится полисифонным в связи с образованием перицентральных клеток. Одна из перицентральных клеток, которая образуется последней, делится, давая несущую клетку и первую клетку карпогонной ветви. Несущая клетка отчленяет еще три стерильные клетки. После оплодотворения карпогон увеличивается в размерах, стерильные клетки делятся, образуя клеточные нити. Несущая клетка отделяет ауксиллярную, которая располагается вблизи от карпогона и сливается с ним посредством маленькой соединительной клетки. В процессе развития гонимобласта несущая, ауксиллярная, центральная и стерильные клетки слоевища сливаются в одну крупную клетку. Карпоспоры конечные. Зрелый гонимобласт окружен клеточной оберткой с отверстием на вершине и имеет шаровидную или грушевидную форму (рис. 187, 6-10). В мужских трихобластах только один базальный и несколько верхушечных сегментов остаются однорядными. Остальные сегменты окружаются перицентральными клетками, которые подвергаются поперечным делениям с образованием трех или более материнских клеток сперматангиев. Те, в свою очередь, делятся косыми перегородками и дают по нескольку сперматангиев (рис. 187, 2-5).
Тетраспорангии возникают на обычных полисифонных ветвях. В каждом сегменте образуется по одному тетраспорангию. Перицентральная клетка отчленяет от своей внешней поверхности три стерильные покровные клетки. Оставшаяся часть этой клетки делится на крупный тетраспорангий и маленькую клеткуножку. Цикл развития полисифонии состоит в правильной смене гаметофитов и тетраспорофита одинакового морфологического строения.
Роды родомела (Bhodomela) и одонталия (Odonthalia) тоже наиболее обычные родомеловые из флоры наших морей (табл. 23, 1, 3). Они относятся к группе родов, у которых перицентральные клетки располагаются в несколько слоев. Кроме того, органы полового размножения образуются у них на специальных коротких полисифонных веточках - стихидиях. Тетраспорангии также возникают в специальных веточках. Стерильные трихобласты (если имеются) пигментированы и не опадают.
Как уже упоминалось, родомеловые берут начало от каких-то примитивных представителей семейства церамиевых и представляют собой самостоятельную, независимую от остальных семейств линию эволюции.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'