Порядок гигартиновые (Gigartinales)
Гигартиновые - чрезвычайно разнообразная группа с нечеткими границами. Это разнообразие проявляется во внешнем и внутреннем строении, в строении органов размножения, в особенностях развития гонимобласта. Уже само количество входящих в порядок семейств - а их насчитывается 21 - говорит о большой его неоднородности. Единственным критерием для выделения порядка служит то, что ауксиллярной клеткой у этих водорослей является обычная клетка слоевища. Однако есть гигартиновые, у которых ауксиллярной клеткой служит несущая, что можно наблюдать и у криптонемиевых. По мере изучения этих водорослей границы между упомянутыми порядками становятся все менее ясными и определение этих границ часто доступно лишь опытным специалистам. Идет постоянный процесс перемещения родов из порядка в порядок. Нередки случаи, когда детали развития оплодотворенного карпогона очень трудно разгадать и объяснить и разные исследователи на одном и том же микроскопическом препарате могут видеть разные вещи. По-видимому, справедливо утверждение о том, что стабильность существующей классификации красных водорослей иллюзорна и что из всех порядков только один - церамиевых - достаточно хорошо отграничен.
Семейство круориевые (Cruoriaceae)
Это семейство представляет как раз пример того, о чем было только что сказано. До недавнего времени эту небольшую группу корковых водорослей помещали в порядок криптонемиевых. По облику круориевые очень напоминают водоросли из семейства сквамариевых, но отличаются структурой. Вертикальные нити, поднимающиеся от гипоталлия, соединены довольно рыхло и легко отделяются друг от друга (рис. 175, 1). Наиболее известны в семействе круория (Cruoria) и петроцелис (Petrocelis).
Размножение круориевых изучено до сих пор довольно плохо. У круории двух -,трехклеточная карпогонная ветвь образуется как веточка ассимиляционной вертикальной нити (рис. 175, 2). После оплодотворения из карпогона вырастают соединительные нити, которые сливаются с обычными вегетативными клетками соседних нитей. Гонимобласт развивается от соединительных нитей. Клетки в верхних частях ассимиляционных нитей отчленяют по одной боковой клетке, которые дают по 2-3 сперматангия (рис. 175, 3). Тетраспорангии, зонально или крестообразно разделенные, образуются из клеток вертикальных нитей интеркалярно или как одноклеточные боковые веточки этих нитей. Для многих круориевых известно только бесполое размножение.
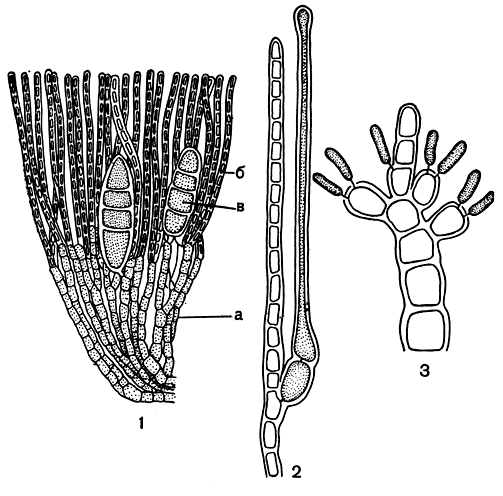
Рис. 175. Строение и размножение Cruoria: 1 - вертикальный срез через корку; 2 - карпогонная ветвь; 3 -веточка со сперматангиями. а - базальный слой, б - вертикальные нити, в - тетраспорангии
У водорослей остальных 20 семейств гигартиновых гонимобласт начинает развиваться из ауксиллярной клетки, а не от соединительных нитей, как у круориевых. Все они делятся на две группы: в первой гонимобласт растет внутрь слоевища, во второй - наружу.
Семейство немастомовые (Nemastomaceae)
Семейство немастомовых включает 4 тепловодных рода водорослей различной формы - от плоской широкопластинчатой до цилиндрической вильчато-разветвленной. Анатомическое строение лучше всего изучено у рода платома (Platoma). Слоевище платомы состоит из уплощенных вильчато- или неправильно разветвленных ветвей. При прорастании спор вначале формируется базальная подушка из плотно соединенных нитей. Из нее вырастают вертикальные побеги. Группы из 4-6 нитей растут вместе, одним пучком, скрепленные слизью (рис. 176, 1). Так образуется центральная ось слоевища фонтанного строения. Пучки боковых ветвей располагаются рыхло и соединяются посредством слизи. От нижних клеток боковых ветвей отходят ризоиды, спускающиеся по слоевищу.
Таким образом, строение немастомовых весьма напоминает строение примитивных представителей порядков немалиевых и криптонемиевых (рис. 176, 2).
По мере усложнения организации происходит более плотное соединение коровых клеток, так что, например, у шизимении (Schizymenia) нитчатую структуру коры можно различить с большим трудом. Центральная часть слоевища шизимении построена из тонких рыхлых нитей, что хорошо видно на поперечном срезе. Широкопластинчатое слоевище ее растет в результате деятельности краевой меристемы (рис. 176, 3).
Каковы же особенности размножения немастомовых? У платомы карпогонная ветвь возникает на одной из нижних клеток ветвей укороченного роста. Соединительные нити вырастают прямо из карпогона и сильно вытягиваются в длину, пока не соединятся с ауксиллярной клеткой, которой служит одна из клеток соседних ветвей ограниченного роста. Первая клетка гонимобласта отчленяется к поверхности слоевища и дает густой пучок нитей, большинство клеток которых превращается в карпоспоры. Гонимобласты не имеют обертки и лежат между ассимиляционными нитями (рис. 176, 4). Зонально разделенные тетраспорангии разбросаны по слоевищу. Они образуются как одноклеточные боковые веточки коровых нитей.
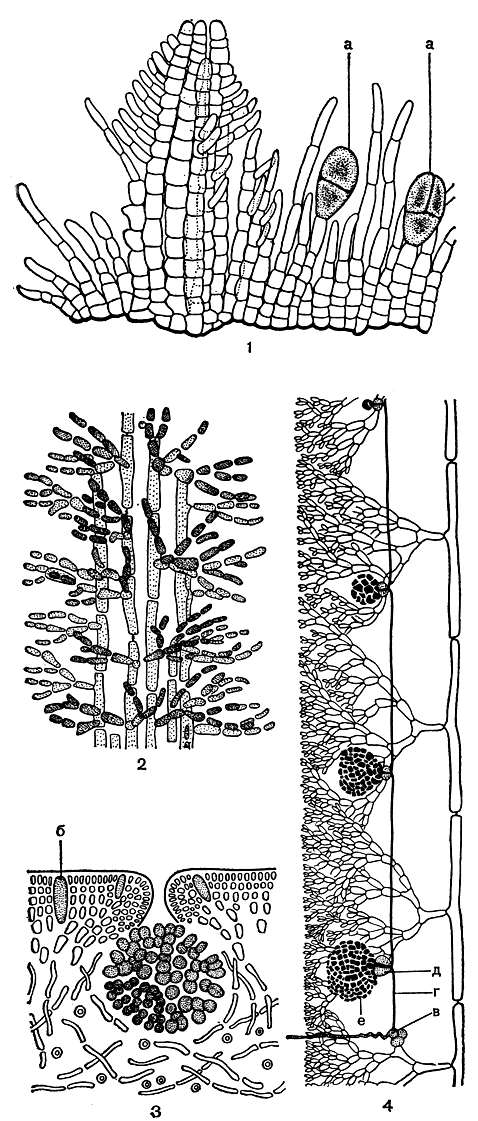
Рис. 176. Строение и размножение немастомовых: 1 - ювенильная стадия Platoma; 2 - часть молодого вертикального побега; 3 - поперечный срез слоевища Schizymenia со зрелым гонимобластом; 4 - развитие гонимобласта у Platoma. а - тетраспорангии, б - железистые клетки, в - карпогонная ветвь, г - соединительная нить, д - ауксиллярная клетка, е - гонимобласт
Семейство филлофоровые (Phyllophoraceae)
Семейство филлофоровых интересно с разных точек зрения. С одной стороны, в него входят такие полезные водоросли, как филлофора и анфельция, промысел которых ведется у нас в Черном, Белом и дальневосточных морях. С другой - некоторым филлофоровым свойственны отклонения от нормального цикла развития, изучение которых важно для решения проблемы циклов развития у красных водорослей. Семейство невелико по объему, оно объединяет 8 родов, из которых заслуживают упоминания гимногонгрус (Gymnogongrus), филлофора (Phyllophora, рис. 177, 1-3) и анфельция (Ahnfeltia, рис. 177, 4).
Филлофора распространена почти во всех морях СССР. В Черном море заросли ее, известные под названием "филлофорное поле Зернова", достигают промысловых размеров. Слоевище филлофоры небольшое, пластинчатое, линейной, клиновидной или овальной формы, разветвленное дихотомически, пальчато или неправильно. Внутренняя часть слоевища, сложенная крупными бесцветными клетками, ограничена тонким коровым слоем. Кора состоит из мелких плотно соединенных клеток, расположенных вертикальными рядами, по 1-4 в ряд (рис. 177, 2).
Органы размножения обычно располагаются на специальных листочках. Ряд видов филлофоры характеризуется полным циклом развития, в котором представлены все формы - гаметофиты, карпоспорофит и свободноживущий тетраспорофит. У филлофоры перепончатолистной (Phyllophora membranifolia) половые растения двудомные. При образовании сперматангиев из поверхностных клеток возникают веточки, на которых, в свою очередь, образуются сперматангии. При этом более длинные плодоносные веточки чередуются с более короткими, и вследствие этого на поверхности слоевища, под внешней оболочкой, образуются своего рода углубления, дно которых выложено сперматангиями. Благодаря такому способу образования сперматангиев поверхность мужских листочков становится мелкоячеистой (рис. 177, 3).
Трехклеточная карпогонная ветвь сидит на клетке, которая является и ауксиллярной (рис. 177, 2). Нити гонимобласта начинают расти внутрь слоевища среди клеток особой питающей ткани, которая возникает еще до оплодотворения. Большинство клеток гонимобласта превращается в карпоспоры, выходящие наружу в результате разрушения стенки цистокарпа, образованной толстой многорядной корой. Сорусы тетраспорангиев расположены в нематециях, имеющих форму подушечек, на свободноживущих спорофитах.
Но есть виды филлофоры, у которых цикл развития проходит иначе. На женских гаметофитах после оплодотворения карпогона развиваются шаровидные нематеции, в которых образуются тетраспоры. Долгое время эти нематеции принимали за другую, паразитическую водоросль. По новейшим данным, это явление можно объяснить следующим образом. По-видимому, здесь карпоспорофит редуцируется до зиготы, которая непосредственно превращается в карпоспору. Карпоспора дает начало тетраспорофиту, который также морфологически редуцирован и развивается на гаметофите, как паразит. Сходные явления наблюдаются в цикле развития близкого к филлофоре рода гимногонгрус. По сравнению с филлофорой этот род относится к более тепловодным организмам и поэтому распространен южнее. Только три вида гимногонгруса (из 40 известных в настоящее время) заходят в самые южные моря СССР - Черное и Японское.
Род анфелъция (табл. 39, рис. 177, 4) распространен в северных частях Атлантического и Тихого океанов и заходит в Арктику. Анфелъция - главное сырье агаровой промышленности в СССР. Ее добывают в Белом море и у побережья Сахалина. Для анфельции очень характерна неприкрепленная форма роста, которая как раз и образует промысловые скопления. В таких скоплениях анфельция размножается только вегетативным путем. У прикрепленных видов, кроме вегетативного, известно размножение моноспорами, которые развиваются в нематециях. Способность к половому размножению утеряна у анфельции полностью.
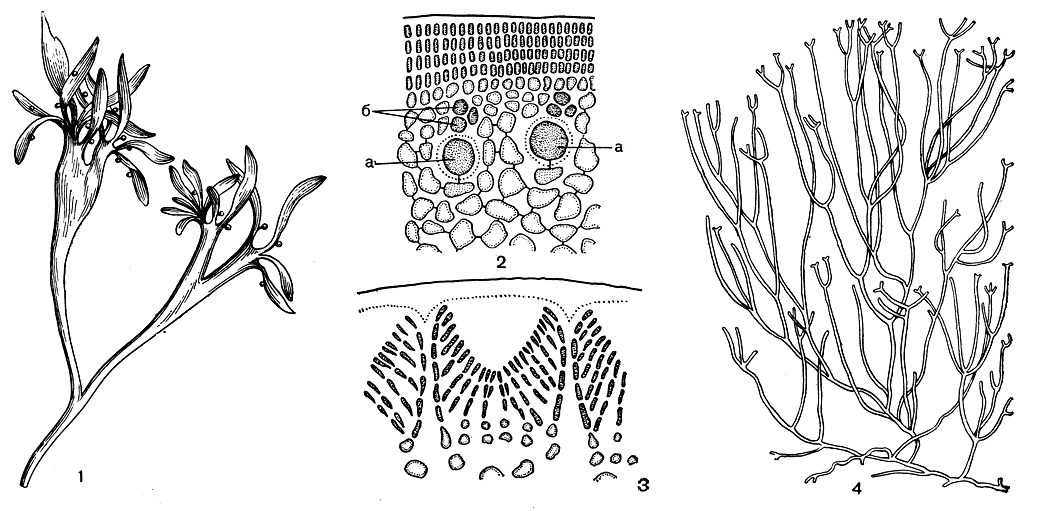
Рис. 177. Строение и размножение филлофоровых: 1 - растение Phyllophora с нематециями; 2 - поперечный срез слоевища Phyllophora; 3 - сперматангии на поперечном срезе; 4 - растение Ahnfeltia. а - ауксиллярная клетка, б - карпогонная ветвь
Семейство гигартиновые (Gigartinaceae)
Небольшое семейство гигартиновых включает пять родов, из них в наших морях обитают четыре - гигартина (Gigartina), хондрус (Chondrus), иридея (Iridea) и родоглоссум (Rhodoglossum). Все они обладают сходным строением, но различаются особенностями органов размножения. Слоевище гигартиновых может быть цилиндрическим, сдавленным или пластинчатым, разветвленным или рассеченным (табл. 20, 4; 21, 2). Характерной формой ветвления является вильчатое; нередко все ветви покрыты пролификациями. В отличие от филлофоровых структура слоевища гигартиновых отчетливо нитчатая. Это хорошо видно на поперечном срезе через слоевище любой водоросли семейства. В центральной части и во внутренней коре тонкие нити рыхло переплетаются наподобие сети. Внешняя кора построена более плотно, но и в ней нетрудно проследить мелкоклеточные дихотомически-разветвленные нити (рис. 178, 2).
Крестообразно разделенные тетраспорангии развиваются группами. У хондруса и иридеи они возникают на клетках центральной части слоевища как короткие разветвленные нити, все клетки которых при созревании превращаются в тетраспоры (рис. 158, 2). У гигартины и родоглоссума тетраспорангии образуются из клеток внутренней части коры. В этом случае они располагаются цепочками, так как в тетраспорангии превращаются несколько клеток каждой коровой нити. Таким образом, если у последних двух родов тетраспорангии соответствуют обычным вегетативным клеткам, то у первых - это дополнительные образования. Характер развития тетраспорангиев - важный, часто единственный признак, помогающий отличить друг от друга роды, во всем остальном, включая анатомическое строение и внешнюю форму, чрезвычайно сходные, например родоглоссум от иридеи.
Строение женской половой системы и развитие цистокарпов сходно у всех членов семейства. Крупная несущая клетка располагается во внутренней коре (рис. 178, 2). После оплодотворения она сливается с карпогоном и дает начало гонимобласту (рис. 178, 3). Зрелые го нимобласты - шаровидные образования, погруженные в слоевище или выступающие над его поверхностью (рис. 178, 4). Обычно они разбросаны по всему телу водоросли, только у гигартины их образование приурочено к специальным бородавчатым выростам на поверхности. По этому признаку род гигартина хорошо отличается от остальных водорослей семейства. У хондруса гонимобласт не имеет специальной оболочки. У остальных родов образование гонимобласта связано с особой питающей тканью. От нее вырастают длинные узкие нити, которые окружают зрелый гонимобласт, образуя его обертку.
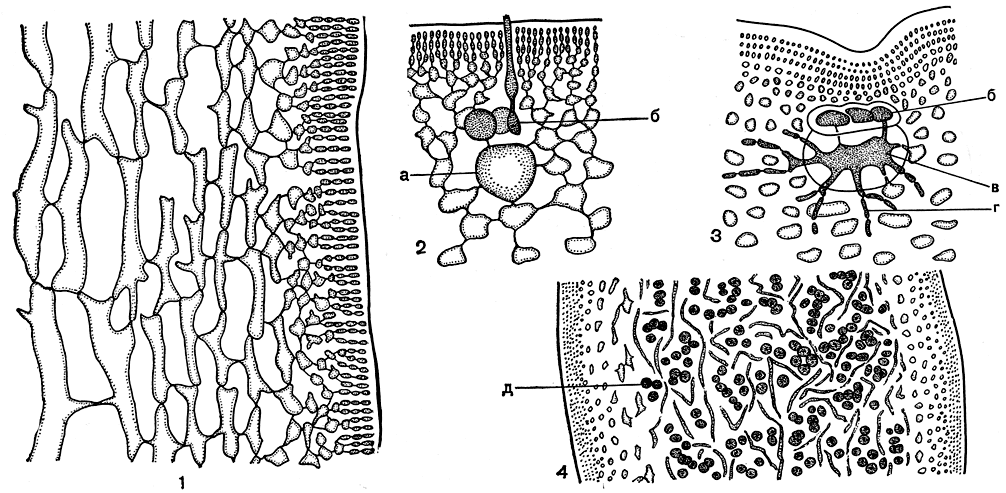
Рис. 178. Строение и размножение гигартиновых: 1 - продольный срез слоевища Chondrus; 2 - карпогонная ветвь; 3 - начало образования нитей гонимобласта; 4 - зрелый гонимобласт на поперечном срезе, а - несущая клетка, б - карпогон, в - клетка слияния, г - нити гонимобласта, д - карпоспоры
Семейство грациляриевые (Gracilariaceae)
Семейство грациляриевых включает несколько небольших родов и один весьма крупный - грацилярия (Gracilaria), более 100 видов которого широко распространены в уме ренных и тропических морях земного шара. Виды грацилярии, как, впрочем, вообще все водоросли порядка, имеют тенденцию уплощаться, поэтому, кроме цилиндрических, есть немало форм с плоским, вплоть до листовидного, слоевищем. Виды грацилярии содержат значительные количества фикоколлоидов и в некоторых странах служат сырьем для агаровой промышленности. Благодаря большому содержанию фикоколлоидов растения грацилярии жесткие, хрящеватые. Взрослое слоевище имеет "клеточную" структуру, каких-либо следов нитчатого строения проследить не удается. Центральная часть состоит из крупных клеток, кнаружи постепенно уменьшающихся в размерах, кора построена из нескольких слоев мелких клеток (рис. 179, 1). Рост слоевища происходит за счет меристемы, которая сформировалась на основе одноосевого строения. Правда, центральную ось не удается проследить даже у проростков, видна только одна апикальная клетка, отчленяющая вниз сегменты от трех своих поверхностей. Особенности размножения состоят в том, что ауксиллярная клетка до оплодотворения неразличима. Двухклеточная карпогонная ветвь образуется на клетках внешней коры (рис. 179, 3). После оплодотворения карпогон сливается с несколькими соседними клетками, образуя массивную клетку слияния, из которой вырастают нити гонимобласта, направленные к поверхности слоевища. После оплодотворения в месте образования прокарпа поверхностные клетки усиленно делятся, образуя толстую мелкоклеточную кору, из которой позднее формируется крыша цистокарпа с отверстием посередине. Цистокарпы, разбросанные по слоевищу, выступают над его поверхностью в виде мелких шариков. Питание развивающегося гонимобласта осуществляется клеткой слияния и отчасти близлежащими клетками слоевища (рис. 179, 4, 5).
Сперматангии у грацилярии образуются в специальных углублениях коры, напоминающих концептакулы, с отверстием на вершине (рис. 179, 2).
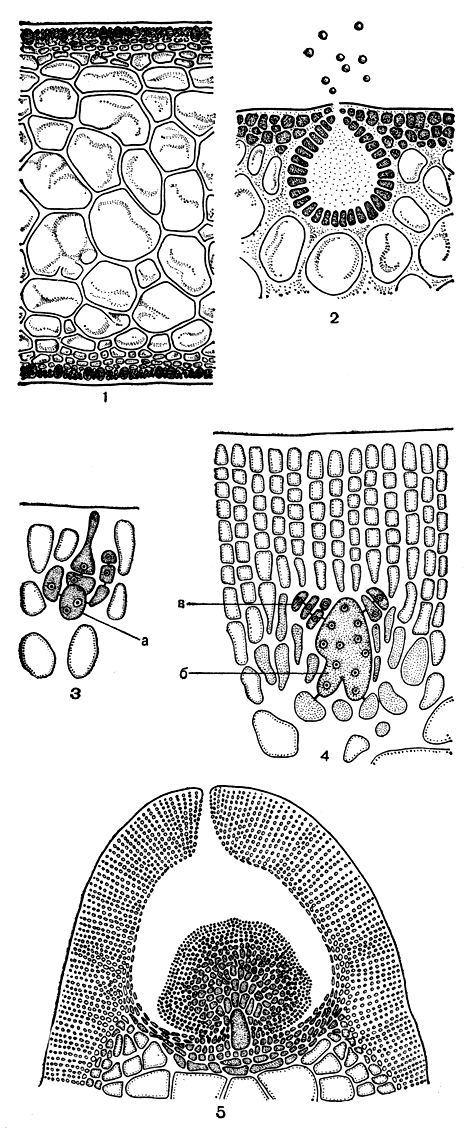
Рис. 179. Строение и размножение Gracilaria: 1 - продольный срез слоевища; 2 - часть среза со сперматангиями; 3 - карпогонная ветвь; 4 - начало развития гонимобласта; 5 - зрелый цистокарп. а - несущая клетка, б - клетка слияния, в - клетки гонимобласта
Крестообразно разделенные тетраспорангии разбросаны в коровом слое по всему слоевищу.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'