4. Происхождение женского гаметофита цветковых растений
У простейших размножение состоит в делении клеток; бактерии и циановые водоросли в короткое время могут образовать значительные массы исключительно путем простого деления. Бактерии клёка за 12 часов нередко превращали на сахарных заводах содержимое чанов с 500 кг патоки в студенистые комья. Клетки их обычно все одинаковы; там, однако, где холод или засуха постепенно прекращают рост организмов, отдельные клетки превращаются в цисты, или покоящиеся споры. При этом оболочка клеток утолщается, происходит накопление в протоплазме запасных питательных веществ и клетка несколько разрастается; она переходит затем в покоящееся состояние и лишь при наступлении благоприятных условий снова начинает ассимилировать, поглощать растворы и делиться.
У жгутиковых организмов, причисляемых к растениям, в некоторых случаях образуются одинаковые между собой гаметы или изогаметы. Это клеточки, снабженные по большей части двумя ресничками, которые, двигаясь в воде, сталкиваются между собой и, теряя реснички, сливаются вместе попарно. Образовавшаяся двухъядерная клетка зигота также превращается в цисту и переживает период покоя, после которого в ней происходит редукционное деление и образуются молодые одноядерные клетки следующего поколения.
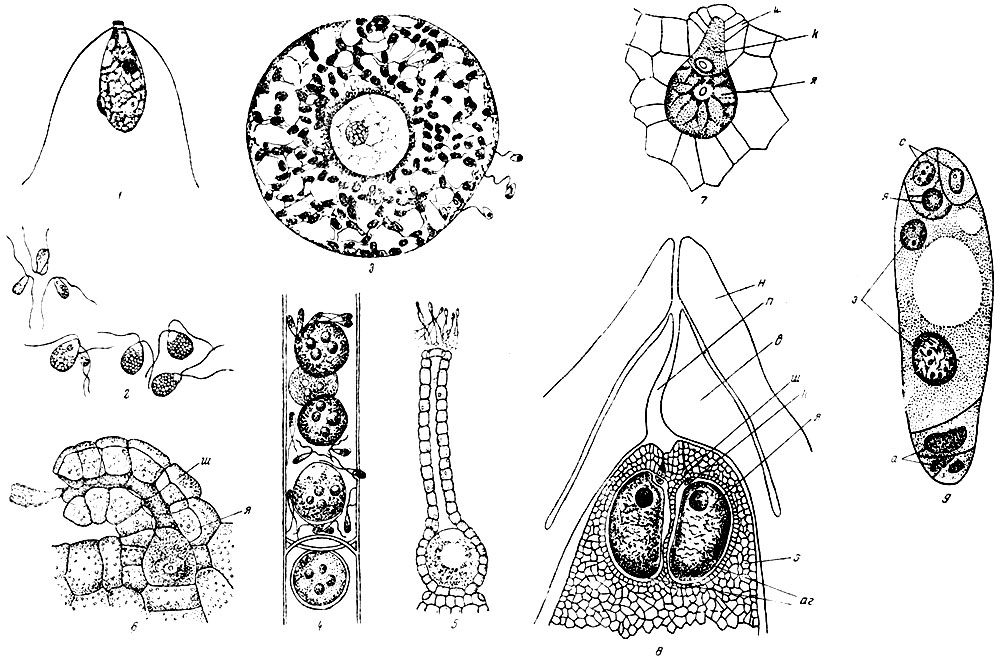
Рис. 26. Развитие гаплоидных элементов от изогамет до зародышевого ядра цветковых растений: 1 - изогамета водоросли 'морокой салат' (Ulva latuca); 2 - гетерогамия: макро- и микрогаметы водоросли Codium; 3 - яйцеклетка водоросли диктиота в момент оплодотворения; справа - микрогаметы (сперматозоиды); 4 - оогоний водоросли (сперматозоиды); 5 - архегоний мха с яйцеклеткой в нижней, расширенной части сфероплеа с несколькими яйцеклетками и проникшими в него извне микрогаметами и сперматозоидами у входа в его шейку наверху; 6 - архегоний папоротника, несколько упрощенный по сравнению с архегонием мха; яйцеклетка частично погружена в ткань заростка. Из канала шейки выходит слизь, получившаяся при расплывании канальцевых клеток; 7 - еще более упрощенный архегоний папоротника сальвиния; 8 - архегонии сосны, находящиеся среди ткани заростка в зародышевом мешке, внутри семяпочки; 9 - зародышевый мешок лилии, вынутый из семяпочки; н - наружный покров семяпочки; п - канал пыльцевой камеры; в - внутренний покров семяпочки; э - ткань заростка, превращенная в эндосперм; аг - два архегония; ш - шейка архегония; к - канальцевая клетка; а - антиподы (ткань заростка); з - ядра зародышевого мешка, из которых после оплодотворения возникает эндосперм; с - спутник или синэргиды; я - яйцеклетка
У водорослей мы имеем уже правильное наличие гамет в цикле развития. На низшей стадии эволюции стоят водоросли. у которых гаметы совершено одинаковы, как, например, у обыкновенной на севере зеленой нитчатки улотрикс, на средней же между гаметами есть уже некоторое различие в размерах - более крупная имеет ресницы и двигается, как это имеет место у простейших водорослей. Крайним случаем такой анизогамии и будет оогамия, так как яйцевая клеточка отличается от изогаметы крупными размерами, обилием запасных питательных веществ и полным отсутствием органов движения - ресниц. Дальнейшим усложнением является все растущая защищенность яйцеклетки водорослей нитчаток, прикрытой лишь оболочкой материнской клетки. У несколько более совершенных форм эта оболочка является в виде специального органа оогония. У многих бурых водорослей оогонии находятся в особых полузамкнутых ямках скафидиях, что значительно увеличивает их шансы на спокойное развитие.
Мы не знаем, как развивались псилофиты, но у современных мхов мы находим в пазухах листьев, часто собранными помногу па концах веточек, еще более сложные органы архегонии. Архегоний состоит из шейки и брюшка, стенка его многоклетная, полость брюшка заполнена крупной мощной яйцеклеткой или иначе макрогаметой; канал шейки выполнен несколькими легко расплывающимися клетками. Проникающий сквозь канал сперматозоид вызывает оплодотворение, связанное, как и всегда, с удвоением числа хромосом. Яйцеклетка превращается в зиготу, последняя делится и образует своим дальнейшим ростом палочкообразный зародыш, из которого позднее образуется спорогон мха.
У мхов, хвощей, настоящих папоротников и некоторых других высших споровых все споры одинаковы, и угадать, вырастет ли из нее гаметофит, способный дать и макро- и микрогаметангии, или же только макрогаметангии - архегонии, или же только микрогаметангии - антеридии, невозможно.
Между тем уже у крупных деревьев девонского и каменноугольного периодов, именно у лепидодендронов, было полное разделение на макро- и микроспоры. Из макроспор вырастали макрогаметофиты, т. е. заростки, дававшие архегонии и в них яйцеклетки. Английскому палеонтологу Макину (Maclean) удалось получить тонкие шлифы через макроспоры, на которых хорошо видно, как образовавшийся уже в макроспоре заросток своей верхней частью выдается из лопнувшей макроспоры, образуя тонкую, вероятно, зеленую волнистую пластинку, на которой сидят архегонии. Большая часть этого заростка, вероятно бесцветная, оставалась внутри споры и состояла из ткани, богатой запасными питательными веществами.
У современных нам плауновых, называемых селагинеллами, у маленькой группы шильников (lsoetaceae), у некоторых водяных папоротников (Salviniaceae и Marsiliaceae) деление на макро- и микроспоры выражено очень хорошо и позволяет во всех деталях проследить развитие мужских и женских заростков. Яйцеклетка у них находится в архегонии, который погружен в ткань заростка. Если бы заросток не выступал из макроспоры, а развивался бы целиком внутри ее, то защита первых стадий зародышевой жизни растения была бы еще более обеспечена, что мы и видим у цветковых растений.
У голосеменных растений редукция (упрощение) гаметофита идет значительно далее. Архегонии состоят из крупной яйцеклетки, па верхушке которой сидит несколько маленьких клеточек, сохранившихся как рудимент шейки архегония.
У голосеменных и цветковых растений, как возникших в процессе борьбы за овладение засушливыми территориями юрских материков, гаметофит и яйцевая клетка глубоко запрятаны под защитой тканей спорофита. Макроспоры возникают путем редукционного деления в глубине тканей семяпочки, в полости завязи, окруженной покровами цветка или соцветия. Из четырех макроспор зрелости достигает только одна, она сильно разрастается и превращается в так называемый зародышевый мешок. крупную клетку с двумя гаплоидными ядрами в центре. Прорастание этой макроспоры и развитие в ней заростка состоят в том, что у верхнего ее конца развиваются три крупных ядра - зародышевое и два его спутника (две синэргиды), а у нижнего три клетки, называемые антиподами. Последние можно рассматривать как вегетативную ткань заростка. Считая и центральные ядра, всего в зародышевом мешке развивается 8 ядер. В отдельных случаях число это значительно больше, так как антипод может быть много.
Заросток с одним архегонием или, как думают другие ученые, с тремя архегониями и тремя яйцеклетками готов. Он совершенно изолирован от влияния подсыхания, тесно связан физиологически с материнским растением, и развитие из яйцеклетки зародыша поставлено в особо благоприятные условия.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'