Препараты родиолы, подобно некоторым другим представителям группы женьшеня, обладают адаптогенными свойствами. Они повышают устойчивость организма к действию различных факторов химической, физической и биологической природы.
По мнению Н. В. Лазарева (1961, 1963), адаптогенное действие лекарственных веществ обусловлено развитием в организме «состояния неспецифически повышенной сопротивляемости» (СНПС). Оно выявляется, как правило, в тех случаях, когда необходимо напряжение компенсаторно-защитных механизмов организма, причем СНПС выражается двояким образом: в виде повышения устойчивости к дополнительным нагрузкам (например, повышение работоспособности) и в виде регулирующего эффекта, симптомом последнего можно считать более быструю нормализацию возникших при различных воздействиях сдвигов, независимо от того, в какую сторону эти отклонения направлены.
Н. А. Минкина и Е. И. Люблина (1968) полагают, что СНПС, возникшее при привыкании к ядам, может проявляться з виде регулирующего влияния, повышения работоспособности и возрастания мышечной силы, увеличения иммунологической активности и сопротивляемости к инфекциям.
СНПС можно достигнуть двумя путями: 1) постепенно приучая организм к воздействию неблагоприятных факторов внешней среды и 2) однократным введением лекарственных веществ, обладающих адаптогенными свойствами (И. И. Брехман, 1968), в частности, психостимуляторов-адаптогенов.
В предыдущих главах было описано повышение препаратами родиолы мышечной работоспособности, что является одним из важных признаков формирования в организме СНПС и обнаруживается при применении различных функциональных нагрузок.
Теперь мы рассмотрим присущие родиоле свойства регулируюшего влияния при некоторых экспериментальных воздействиях, вызванных факторами химической и биологической природы.
Таблица 28. Антинаркозное действие препаратов родиолы и элеутерококка (Р. А. Аксенова, 1968; А. И. Елькин, 1970а)
Препарат
Доза на кг веса
Колич. мышей
Время наступления сна, мин.
Длительность сна, мин.
M±m
P
M±m
P
Опыты с барбитал-натрием (Барбитал-натрий, гексанал и хлоралгидрат вводили подкожно.)(150 мг/л)
Контроль Экстракт родиолы Салидрозид
5 мл 5 мл 100 мг
23 39 35
33±4,5 38±4,6 35±1,8
0,42 0,68
237±25,6 133±22,8 147±10,5
0,002 0,002
Опыты с бензином (В парах бензина мыши находились 60, ацетона - 50, эфира - 5 минут, затем животных извлекали из камеры и фиксировали длительность сна.)(50 мг/л)
Контроль Родозин Экстр. элеутерококка
5 мл 5 мл 5 мл
10 10 10
33±3,7 44±2,8 42±3,1
0,015 0,071
11±3,7 2±0,4 3,2±0,4
0,015 0,035
Опыты с ацетоном (120 мг/л)
Контроль Родозин Экстр. элеутерококка
5 мл 5 мл 5 мл
17 17 17
32±1,1 33±1,0 34±1,3
0,48 0,27
5,0±1,2 6,0±1,2 5,3±0,7
0,54 0,84
Опыты с гексаналом (70 мг/л)
Контроль Родозин Экстр. элеутерококка
5 мл 5 мл 5 мл
10 10 10
9±0,2 16±0,5 15±0,8
0,000 0,000
39±2,3 21±0,6 20±0,9
0,000 0,000
Опыты с хлораогидратом (300 мг/кг)
Контроль Родозин Экстр. элеутерококка
5 мл 5 мл 5 мл
10 10 10
21±1,3 19±1,4 20±1,8
0,31 0,61
19±2,2 20±2,6 19±1,9
0,54 1,0
Опыты с эфиром (В опытах с эфиром время в секундах.)(70 мг/л)
Контроль Родозин Экстр. элеутерококка
5 мл 5 мл 5 мл
10 10 10
118±2,7 151±3,2 145±2,7
0,000 0,000
156±3,6 118±1,6 127±3,7
0,000 0,000
Антитоксическое действие родиолы проявляется по отношению к различным химическим веществам. Как и другие психостимуляторы-адаптогены, она обладает антигипнотическими и антинаркозными свойствами. На табл 28 обобщены результаты исследования антинаркозного действия препаратов родиолы и элеутерококка. Как видно из таблицы, экстракт радиолы (деалкоголизированный), родозин и салидрозид, введенные белым мышам подкожно или внутрибрюшинно за 30—60 минут до воздействия наркотика, ослабляют наркозный эффект (удлиняют время засыпания и сокращают продолжительность сна) бензина, барбитал-натрия, гексенала, эфира и не оказывают пробуждающего действия по отношению к хлоралгидрату и ацетону. Наиболее сильное антинаркозное действие выявлено в опытах с бензином. Так, если длительность сна контрольных мышей при действии наркотика принять за 100%, то у животных, получавших предварительно родозин, она составляла в опытах с бензином 19%, гексеналом и барбитал-натрием — 54 и 62%.
Введение мышам 0,1 мл экстракта родиолы в течение 10 дней увеличивает ЛДво 40%-ного этилового спирта с 24,1 до 56,2 мл/кг (М. Ф. Нехода).
Исследования М. И. Зотовой на 12 добровольцах (мужчины в возрасте 20—28 лет) с помощью корректурного теста по вышеописанной методике (гл. II) выявили положительное влияние экстракта родиолы на умственную деятельность при одновременном назначении со 100 мл 40%-ного спирта. В контрольных наблюдениях через час после приема спирта число прокорректированных знаков существенно не изменилось, но резко возросло количество ошибок (на 77%). У лиц, получавших спирт с добавлением 10 капель экстракта родиолы, число ошибок увеличилось лишь на 15%.
Судя по наблюдениям А. И. Елькина (1973), антинаркотический эффект препаратов родиолы осуществляется при участии М- и Н-холинореактивных систем. На фоне действия холинолитиков — пентамина (100 мг/кг), пахикарпина (50 мг/кг) и атропина (1,6 мг/кг) — антигипнотическое влияние родозина на показатели гексеналового сна либо не проявляется, либо извращается (удлинение продолжительности сна).
Антитоксическое действие родиолы выявлено по отношению к метгемоглобинообразователям: нитриту натрия и анилину (А. И. Елькин, 1970 б, 1971). Внутрибрюшинное введение белым мышам родозина в дозе 5 мл/кг за 30 минут до инъекции нитрита натрия (180 мг/кг) достоверно увеличивало как число выживших животных, так и продолжительность жизни погибших мышей. В контрольной группе после введения нитрита натрия выжило 18% животных, родозин сохранил жизнь 60% мышей. В опытах с анилином (650 мг/кг) существенное увеличение числа выживших животных наблюдалось лишь после профилактического ежедневного введения 2 мл/кг родозина в течение 7 дней. Препарат не влиял на образование и скорость исчезновения из крови телец Гейнца.
Четкий защитный эффект родозина (5 мл/кг) обнаружен при остром отравлении мышей хлорофосом (750 мг/кг подкожно): гибель животных снизилась с 73 до 26% (р<0,001). Родозин не влиял на антихолинэстеразную активность хлорофоса; препарат не вызывал существенных изменений активности холинэстеразы крови и мозга как у интактных мышей, так и у животных, подвергнутых интоксикации хлорофосом (А. И. Елькин, 1973).
Родозин в дозе 5 мл/кг при профилактическом введении белым мышам в течение 7 дней ослаблял интенсивность судорог и статистически значимо (р = 0,001) увеличивал выживаемость животных при действии токсических доз стрихнина (1,0—1,2 мг/кг). Однократная инъекция родозина оказалась неэффективной (Г. С. Хохлов, 1968а).
Получены положительные результаты пои профилактическом введении родозина мышам, находящимся в герметической камере (Г. С. Хохлов, 1968 б). Препарат задерживал развитие судорог и вдвое увеличивал продолжительность жизни подопытных животных (на 19,8 минут; р<0,001). (Экстракт элеутерококка в этих условиях оказался неэффективным, что согласуется с литературными данными (А. А. Константинов, 1965).) По-видимому, этот эффект обусловлен снижением под влиянием родозина потребления кислорода животными. В экспериментах с регистрацией газообмена мышей в приборе закрытого типа родозин через 1 час после введения уменьшал утилизацию кислорода подопытными животными на 10% (с 9,1±0,06 до 8,1 ±0,28 мл/мин/100 г веса тела; р<0,05). Вместе с тем препарат тормозил депримирующее влияние хлорида лития (500 мг/кг внутрибрюшинно) на газообмен (В. М. Шолохов).
Важным свойством веществ, обладающих адаптогенным действием, является их регулирующий, нормализующий эффект, наступающий независимо от направленности предшествующих сдвигов. В отношении препаратов группы женьшеня это свойство впервые было установлено в нашей лаборатории. Экстракты женьшеня, левзеи и элеутерококка препятствуют развитию у кроликов экспериментальных гипер- и гипогликемии, лейкоцитоза и лейкопении, эритроцитоза и эритропении (Р. А. Пичурина. 1963; А. С. Саратиков и Р. А. Пичурина, 1965). (Сходные результаты позднее были получены в лаборатории И. И. Брехмана (И. В. Дардымов. 1965; О. И. Кириллов и И. В. Дардымов, 1966) в опытах с некоторыми гормональными препаратами, являющимися физиологическими антагонистами. Вес надпочечников крыс, увеличивающийся под влиянием АКТГ или уменьшающийся под действием кортизона, при одновременном введении с гормональными препаратами экстракта элеутерококка оставался почти неизменным. Элеутерококк препятствовал гипертрофии (вызываемой введением 6-метилтиоурацила) и атрофии (индуцируемой назначением тиреоидина) щитовидных желез крыс.)
Соответствующие эксперименты (М. И. Зотова, 1965; Р. А. Аксенова, 1968) показали, что аналогичным действием в отношении указанных патологических реакции периферической крови обладают и препараты родиолы.
Опыты проводили на кроликах весом 1,7-3,8 кг. В течение 18 часов до опыта животные голодали. Кровь для исследования брали из краевой вены уха. Лейкоцитоз вызывали однократным введением под кожу спины очищенного скипидара в дозе 0,2 мл/кг. Для воспроизведения лейкопении кроликам внутривенно вводили 0,05 мл/кг дизентерийной вакцины Флекснера «2А», содержащей 1 млрд. микробных тел в 1 мл. Гипергликемию вызывали подкожной инъекцией 0,05 мл/кг 0,1%-ного раствора адреналина, а гипогликемию - введением 1 ЕД/кг инсулина. Исследуемые препараты инъецировали подкожно: салидрозид в дозе 20 мг/кг, экстракт родиолы (деалкоголизированный) — 1 мл/кг.
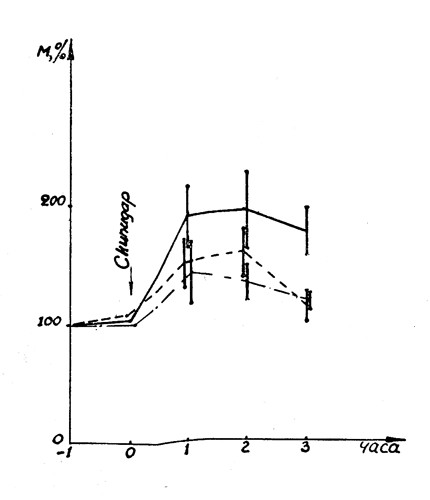
Рис. 21. Влияние салидрозида и экстракта родиолы на развитие лейкоцитарной реакции. Обозначения: ________ контроль, ----- салидрозид, -•-•-•-•- экстракт родиолы.
Введение препаратов родиолы тормозит развитие экспериментального лейкоцитоза (рис. 21). В контроле количество лейкоцитов через 1 час после инъекции скипидара резко возрастало, достигая 183±16,3% но отношению к исходному фону (9200 в 1 мм3) и оставалось на высоком уровне в течение 3 часов опыта. На фоне профилактического введения экстракта родиолы и салидрозида лейкоцитарная реакция выражена значительно слабее (р<0,02—0,001): через 1 час после инъекции скипидара количество лейкоцитов составляло соответственно 136±9,5 и 139±8,3%, а через 3 часа — 118±3,5 и 116±6,6% к исходному фону.
Экстракт родиолы при введении с «лечебной» целью, т. е. на фоне уже развившейся лейкоцитарной реакции (количество лейкоцитов составляет 195% к фону), уменьшал содержание лейкоцитов в периферической крови, однако статистический достоверный эффект наблюдался только через 2 часа (М. И. Зотова, 1965). Вместе с тем препарат предотвращал развитием эндотоксиновой лейкопении. В контрольных опытах через 2 часа после введения вакцины Флекснера «2-А» количество лейкоцитов снижалось на 45%, а на фоне экстракта родиолы лишь на 15%.
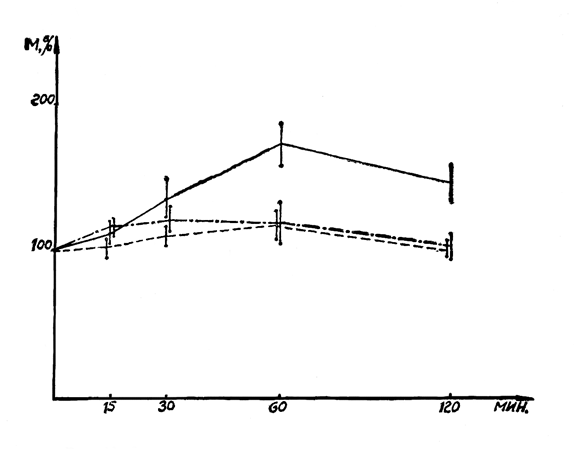
Экстракт родиолы и салидрозид обладают слабым и непродолжительным гипогликемическим действием. Уровень сахара в крови (в среднем 91 мг%) через 30 минут после их введения снижался па 10—14 мг%. Однако оба препарата проявляют выраженный антигипергликемический эффект. Введенные за 15 минут до инъекции адреналина, они препятствуют развитию гипергликемии (рис. 22). Если в контрольных опытах через 60 минут после введения адреналина уровень сахара в крови достигал максимальной величины 150 мг%, то в опытах с экстрактом родиолы и салидрозидом максимальное повышение уровня сахара составляло лишь 117 и 97 мг%.
Клиническая проверка (Л. Ф. Колмакова и Н. И. Кутолина, 1966) выявила слабый гипогликемический эффект у больных, страдающих сахарным диабетом, при назначении им внутрь экстракта родиолы (см. гл. VII).
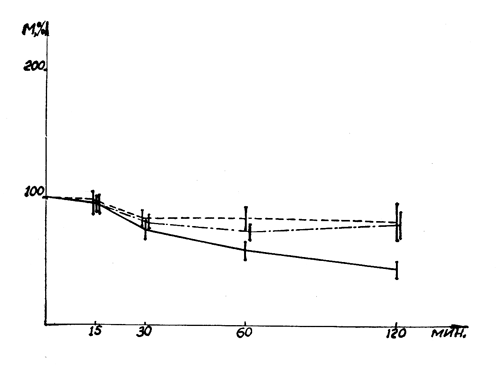
Антигипогликемическое действие родиолы исследовано на фоне тяжелой инсулиновой гипогликемии. Подкожное введение инсулина вызывало у интактных кроликов понижение концентрации сахара в крови, сопровождающееся у части животных судорогами. Исходный уровень сахара (в среднем 104 мг%) к 60 минуте достигал 62, а к 120 минуте — 46 мг%. Профилактическое (за 15 минут) введение экстракта родиолы и салидрозида препятствовало развитию инсулиновой гипогликемии. Степень снижения концентрации сахара в крови не превышала 19% (в контроле 56%) (рис. 23).
В формировании СНПС, по-видимому, участвуют высшие отделы ЦНС и некоторые эндокринные железы. В. Я. Русин (1968) при исследовании механизмов развития СНПС с помощью фармакологических веществ, избирательно блокирующих функцию различных отделов нервной системы, выявил существенную роль в формировании СНПС ретикулярной формации ствола мозга.
Рис. 22. Влияние салидрозида и экстракта родиолы на развитие адреналиновой гипергликемии. Обозначения: ________ контроль, ----- салидрозид, -•-•-•-•- экстракт родиолы.
Хотя имеются существенные различия между СНПС и классическим генерализованным адаптационным синдромом Селье (О. И. Кириллов, 1966), несомненно участие гипофизарно-адреналовой системы в механизме адаптогенного действия препаратов группы женьшеня (М. А. Розин, 1963; И. И. Брехман, 1968). Нарушение целостности различных звеньев нервно-гуморальных механизмов регуляции (повреждение головного мозга, удаление гипофиза, надпочечников, половых желез) резко ослабляет или полностью исключает ряд специфических проявлений действия женьшеня и элеутерококка.
К аналогичному выводу пришли наши сотрудники (С. Г. Чердынцев, М. И. Зотова. 1966: Р. А. Аксенова и соавт., 1968; Б. Ю. Сальник и соавт., 1968), исследуя участие больших полушарий головного мозга, гипофиза, надпочечников, половых желез в реализации ряда эффектов препаратов родиолы (см. также гл. V).
Рис. 23. Влияние салидрозида и экстракта родиолы на развитие инсулиновой гипогликемии. Обозначения: Рис. 22. Влияние салидрозида и экстракта родиолы на развитие адреналиновой гипергликемии. Обозначения: ________ контроль, ----- салидрозид, -•-•-•-•- экстракт родиолы.
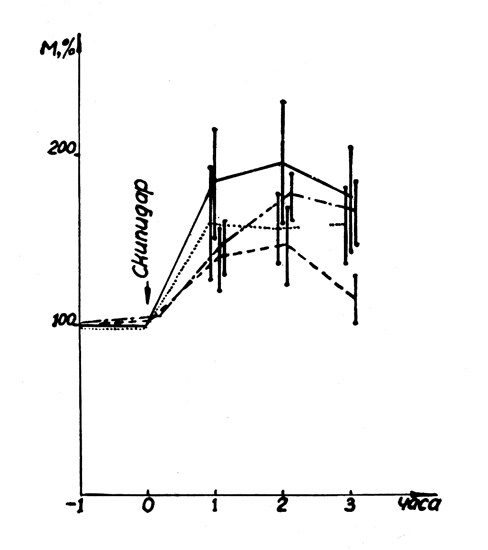
Для анализа антилейкоцитарного действия салидрозида (Р. А. Аксенова, 1968) последний в дозе 20 мг/кг вводили кроликам через 10—12 дней после удаления полушарий головного мозга, гипофизэктомии или кастрации. Предварительные опыты показали, что само удаление полушарий головного мозга у кроликов существенно не отражается на характере лейкоцитарной реакции, наступающей после введения скипидара, по сравнению с интактными животными. Через час после инъекции скипидара количество лейкоцитов в крови контрольной группы бесполушарных животных достигало 160±15% от исходного уровня и оставалось повышенным в течение трех часов наблюдения. Профилактическое введение салидрозида кроликам с удаленными полушариями головного мозга не препятствовало развитию «скипидарного» лейкоцитоза, хотя у интактных животных антилейкоцитарный эффект четко выражен (рис. 24).
Лейкоцитарная реакция на введение скипидара у гипофиз-эктомированных кроликов развивается медленнее, чем у интактных животных и достигает максимальной интенсивности (207±20%) лишь к третьему часу. Салидрозид существенно не изменяет развитие лейкоцитоза у животных с удаленным гипофизом. Сходные результаты получены и в опытах на кастрированных животных.
С целью выяснения роли гормонального фона в механизме антилейкоцитарного действия салидрозида были проведены наблюдения на кастрированных кроликах-самцах, получавших гормонозаместительную терапию, начиная со второго дня послеоперационного периода. Тестостеронпропионат (0,5 мг/кг) в масляном растворе вводили подкожно через день в течение 10 дней.
У кастрированных животных, которым проводилась гормонотерапия, профилактическое введение салидрозида статистически существенно (р<0,001) тормозило развитие лейкоцитарной реакции: через 3 часа после инъекции скипидара количество лейкоцитов составляло 112±7% (в контроле — 159±8%).
Рис. 24. Влияние салидрозида на развитие лейкоцитарной реакции. Обозначения: Рис. 23. Влияние салидрозида и экстракта родиолы на развитие инсулиновой гипогликемии. Обозначения: Рис. 22. Влияние салидрозида и экстракта родиолы на развитие адреналиновой гипергликемии. Обозначения: ________ контроль, ----- салидрозид; бесполушарные животные: ....... контроль, -•-•-•-•- салидрозид.
Таким образом, для реализации адаптогенных свойств родиолы необходимо наличие в организме определенного гормонального фона. По-видимому, этот эффект объясняется свойством некоторых гормонов проявлять так называемые пермессивное действие (Ingle, 1944). Наличие небольших количеств гормонов необходимо для осуществления некоторых реакций организма, непосредственно не связанных с эндокринной системой, они выполняют роль как бы разрешающего фактора.
Анализ антигипер- и антигипогликемических эффектов салидрозида и экстракта родиолы показал, что эти препараты существенно не изменяют характер гипергликемической реакции на подкожное введение адреналина и гипогликемии после инъекции инсулина у кроликов с удаленными полушариями головного мозга, а также после гипофизэктомии или кастрации. Сами по себе эти процедуры (удаление полушарий, гипофиза, половых желез) существенно не отражаются на развитии гипергликемии и гипогликемии.
Необходимость целостности нейрогормональной регуляции в осуществлении метаболических эффектов препаратов родиолы и элеутерококка установили Б. Ю. Сальник (1970) и С. Г. Чердынцев (1970), которые исследовали влияние родозина и экстракта элеутерококка на некоторые показатели энергетического обмена крыс после частичного удаления больших полушарий головного мозга или адреналэктомии.
Как видно из табл. 29, введение родозина и экстракта элеутерококка интактным животным в состоянии покоя сопровождается повышением концентрации сахара в крови; содержание креатинфосфата и гликогена в скелетных мышцах существенно не изменяется. На фоне повреждения полушарий мозга и адреналэктомии эти препараты не вызывают значительных изменений исследуемых показателей.
Поскольку действие родиолы и элеутерококка на энергетический метаболизм проявляется преимущественно на фоне утомления, дальнейшие наблюдения были проведены на животных, подвергнутых воздействию физической нагрузки. В качестве последней использовали плавание крыс в течение 2 часов, так как более длительное плавание приводило крыс к гибели.
Как у интактных животных, так и у крыс с частично удаленными полушариями после 2-часовой работы наступало статистически достоверное снижение концентрации сахара в крови, а также КФ и гликогена в мышцах. У адреналэктомированных животных значительно снижалось лишь содержание гликогена в мышцах.
Существенно различным оказалось влияние психостимуляторов на исследуемые показатели у интактных животных и в условиях экспериментальной патологии. Так у интактных животных экстракт элеутерококка и особенно родозин препятствовали снижению в мышцах под влиянием 2-часового плавания содержания КФ и гликогена, тогда как в опытах на крысах с поврежденными полушариями мозга или удаленными надпочечниками их нормализующий эффект не проявлялся.
Таблица 29. Влияние родозина и экстракта элеутерококка на некоторые показатели углеводно-фосфорного обмена (в мг%) крыс после частичного удаления полушарий головного мозга или адреналэктомии (средние из 8-10 наблюдений).
Показатели
Контроль
Родозин
Экстракт элеутерококка
покой
2 час. плавания
покой
2 час. плавания
покой
2 час. плавания
Интактные животные
Сахар крови Рф Рк КФ мышц Рф Рк Гликоген мышц Рф Рк
Таким образом, повреждение головного мозга и адреналэктомия блокируют проявление адаптогенного действия родозина и экстракта элеутерококка на энергетический метаболизм как в состоянии физиологического покоя, так и особенно при утомлении. Аналогичную направленность изменений наблюдала Л. И. Ямпольская (1952) при угнетении ЦНС тренированных животных амиталом: наркоз, не вызывая существенных изменений в содержании КФ и гликогена мышц, повысившегося под влиянием тренировки, устранял специфику использования источников энергии при выполнении стандартной работы.
Эксперименты с гормонозаместительной терапией у адреналэктомированных крыс (введение ацетата гидрокортизона внутримышечно один раз в сутки по 2,5 мг/100 г, начиная со второго дня послеоперационного периода) свидетельствуют о несомненной роли гормонального фона в механизме действия препаратов родиолы. У адреналэктомированных животных, подвергнутых воздействию физической нагрузки, на фоне гормонозаместительной терапии наблюдается восстановление нормализующего действия родозина на концентрацию КФ в скелетной мышце.
Защитное действие родиолы, как, очевидно, и других психостимуляторов-адаптогенов (А. С. Саратиков, Р. А. Пичурина, 1965; И. И. Брехман, 1968), не является универсальным и несомненно обладает определенной избирательностью, обусловливающей степень эффективности того или другого адаптогена при различных патологических состояниях.
Препараты родиолы (экстракт и родозин по 1 мл/кг), добавляемые в пищу кроликов вместе с холестерином (0,3 г/кг) в течение 4 месяцев, судя по результатам макро- и микроскопического исследования (Д. А. Грацианов, В. Б. Прикс, 1968), не тормозят развитие экспериментального холестеринового атеросклероза и не препятствуют продолжающемуся нарастанию атерогенеза по прекращении введения холестерина.
* * *
Важным и вместе с тем весьма дискуссионным является вопрос о влиянии адаптогенов на течение инфекционного процесса и особенно на неспецифическую иммунобиологическую реактивность организма. Как известно, неспецифическая реактивность находит свое отражение в реакциях организма на самые различные раздражители и связана с его генетическими и физиологическими свойствами. Факторы неспецифической иммунологической реактивности обеспечивают повседневную защиту организма от многочисленных микробов внешней среды, а в случае возникновения болезни способствуют его защитным силам справиться с инфекцией. В биологическом аппарате защиты организма различают гуморальные факторы (комплемент, пропердин, лизоцим и др.) и клеточные. Их совокупность и создает барьер для развития инфекционного процесса.
Н. В. Лазарев (1961, 1963) считает повышение устойчивости к инфекциям одним из характерных свойств СНПС, отличающим это состояние от «стадии тревоги» генерализованного адаптационного синдрома, при котором резистентность организма к инфекционному процессу понижена (Г. Селье, 1960). По мнению Н. В. Лазарева, СНПС наступает, минуя «стадию тревоги» с присущим ей отягощением течения острой инфекции.
Действительно, П. П. Голиков и Н. П. Иконников (1962) сообщили о значительной эффективности в отношении сезонных катаров верхних дыхательных путей профилактического (в течение месяца) назначения экстракта элеутерококка (по 0,5 мл на прием) 180 молодым мужчинам в процессе их акклиматизации к условиям Приморского края.
Ю. И. Бронников (1966) описал благоприятное влияние элеутерококка на резистентность мышей к заражению культурой дизентерийных бактерий Флекснера. По нашему мнению, этот вывод следует принять с осторожностью, поскольку мыши обладают высокой естественной резистентностью к данной инфекции и начинают быстро разрушать дизентерийные бактерии с освобождением эндотоксина. Отмеченное автором увеличение выживаемости животных под влиянием элеутерококка, по-видимому, объясняется антитоксическим действием препарата в отношении эндотоксина. С этим предположением согласуются результаты исследований Н. Н. Самойлова (1966, 1967, 1968), в опытах которого введение экстракта элеутерококка внутрь не отражалось на течении паратифозной инфекции мышей. При внутрибрюшинной инфекции препарат отягощал течение болезни и сокращал продолжительность жизни зараженных животных. У мышей, подвергшихся воздействию элеутерококка, обсемененность печени и селезенки палочкой Бреслау была более выражена, чем у контрольных животных.
И. И. Брехман (1968) на основании экспериментов своих сотрудников (М. И. Положенцева, 1964; Е. Г. Ливкина, А. И. Соловьева, 1965; М. И. Положенцева и Т. Л. Быховцова, 1966; И. В. Дардымов и соавт., 1966) считает, что элеутерококк стимулирует выработку в организме антител (иммунизация брюшнотифозной вакциной и салмонеллами мышиного тифа) и благоприятно влияет на общую иммунологическую реактивность организма.
Кафедрой микробиологии Томского мединститута (зав. академик АМН СССР проф. С. П. Карпов) и Томским НИИВС совместно с нами проведены детальные исследования влияния родознна и экстракта элеутерококка на развитие инфекционного процесса и иммунобиологическую реактивность организма.
В качестве модели острого инфекционного процесса использовали экспериментальный листериоз у белых мышей и кроликов, сходный по патогенезу и клинике с заболеванием, встречающимся в естественных условиях как у животных, так и у людей (Г. В. Черкашин, 1966,1967,1968). Животных заражали внутривенным (кролики) или внутримышечным (мыши) введением суточной культуры листерий штамма 15/57 1 серотипа. Спустя 12—24 часа развивалось острое септическое заболевание с преимущественным поражением паренхиматозных органов, острый период которого заканчивался в случаях благоприятного исхода к 8—10 суткам.
Подкожное введение родозина (1 мл/кг кроликам и 5 мл/кг мышам) одновременно с заражением и затем ежесуточно после него в течение 7 дней, а также профилактическое введение препарата на протяжении 15 дней перед заражением повышало резистентность животных к листерийной инфекции. Так, ЛД50 для мышей, получавших родозин, была в 1,5—2 раза больше, чем в контрольной группе (3,5 млн. в опыте и 1,6 млн. микробных тел в контроле). У кроликов также наблюдалось более легкое течение инфекции. Животные, которым вводили родозин, меньше теряли в весе и быстрее его восстанавливали, лихорадочная реакция у них была более кратковременной, «лимфоцитарная оздоровительная фаза» наступала раньше и была более выражена, титр комплемента на высоте инфекции был выше, а титр специфических антител ниже, чем в контроле; значительно снижалась гибель животных — с 62,5 до 9,1%.
Изучение распространяемости листерий, введенных мышам под плантарный апоневроз в дозе 200 млн., выявило через 2 и 6 часов меньшую обсемененность органов у животных, получавших родозин.
Заслуживает внимания зависимость эффекта родозина от стадии инфекционного процесса. Введение препарата мышам через 12 часов после заражения, т. е. к концу периода инкубации, значительно снижало резистентность животных к инфекции, тогда как инъекция той же дозы родозина спустя сутки после заражения, напротив, в 2 раза уменьшала процент гибели мышей.
Наблюдаемое Г. В. Черкашиным облегчение течения листерийной инфекции у животных под влиянием родозина, по-видимому, является проявлением специфических свойств этого препарата (По неопубликованным данным Н. Б. Сидоренковой (Алтайский мединститут, 1961), настой родиолы в опытах т уНго обладает бактериостатическим действием в отношении стрептококка и стафилококка.) и не отражает направленность действия других представителей психостимуляторов-адаптогенов группы женьшеня. Действительно, исследование по аналогичной схеме эффективности экстракта элеутерококка показало, что подкожное введение этого препарата (1 мл/кг кроликам и 5 мл/кг мышам) одновременно с заражением листериями и затем ежесуточно на протяжении 7 дней после него отягощает течение и ухудшает исход острой листерийной инфекции у мышей и кроликов. Под влиянием препарата в 2 раза уменьшается ЛД50 для мышей. У кроликов во время инфекции происходит большая потеря веса, лихорадочный период удлиняется, запаздывает подъем комплемента в период становления инфекции и в дальнейшем он находится на низком уровне, лейкоцитоз более выражен, а «лимфоцитарная оздоровительная фаза» задерживается, титры специфических антител выше, чем у контрольных животных. Следует отметить, что низкие титры комплемента при многих инфекционных заболеваниях, в том числе при экспериментальном листериозе (А. Б. Ласинскайте, 1965; Ю. Н. Одинцов, 1965), свидетельствуют о неблагоприятном характере их течения. Очевидно, и более высокие титры специфических антител у кроликов, получавших элеутерококк, следует объяснить тяжестью инфекционного процесса. Известно, что высота титров специфических антител при многих бактериальных инфекционных заболеваниях в большей степени обусловлена не напряженностью иммунитета, а остротой процесса и скоростью освобождения организма от возбудителя, обусловливающего постоянное антигенное воздействие на него (К. Н. Лобан и Е. П.Савицкая, 1956; П. А. Вершилова и соавт. 1965).
Таким образом, как однократное, так и многократное введение элеутерококка животным, зараженным листериями, вызывает утяжеление инфекционного процесса.
Иная картина наблюдается при профилактическом использовании элеутерококка. Предварительное введение препарата в течение 15 дней до заражения листериями несколько облегчает течение заболевания. У кроликов наблюдалась меньшая потеря веса, более короткий лихорадочный период, ускорение наступления «лимфоцитарной оздоровительной фазы» со стороны крови; титр комплемента на высоте инфекции и титр специфических антител существенно не отличались от показателей контрольной группы. Профилактическое введение элеутерококка не оказало существенного влияния на специфическую аллергическую реакцию лапок сенсибилизированных мышей, не менялась также и их неспецифическая воспалительная реакция в ответ на введение микробной массы листерий.
На фоне иммунизации кроликов живой бруцеллезной вакциной родозин при курсовом введении вызывал увеличение количества лейкоцитов, титров комплемента и агглютининов; препарат препятствовал снижению фагоцитарной активности лейкоцитов по отношению к листериям (А. А. Триполитова и соавт., 1966). В то же время родозин в процессе иммунизации кроликов столбнячным анатоксином не оказывал существенного влияния на комплементарную и лизоцимную активность крови, гематологические показатели (РОЭ, содержание гемоглобина, количество эритроцитов и лейкоцитов, общий белок и белковые фракции сыворотки), незначительно стимулировал выработку столбнячного антитоксина (увеличение титра в 1,8 раза по сравнению с контролем, р<0,001) (Л. П. Сагайдак, О. Г. Пазникова, 1966).
Экстракты родиолы и левзеи существенно не влияют на формирование у морских свинок специфического иммунитета к вирусу клещевого энцефалита (реакции биологической нейтрализации, торможения гемагглютинации и связывания комплемента), тогда как элеутерококк несколько повышает титр противовирусных антител. Отмечено резкое усиление плазмо-цитарной реакции в ответ на введение указанных препаратов в опытах с иммунизацией морских свинок антигеном вируса клещевого энцефалита (Ю. В. Федоров и соавт., 1966).
Для решения вопроса о целесообразности использования психостимуляторов как средств повышения иммунобиологической сопротивляемости организма Е. П. Красноженов (1968, 1969, 1970) исследовал неспецифическое звено иммунитета при однократном и курсовом введении пиридрола, родозина и экстракта элеутерококка интактным животным и на фоне снижения реактивности с помощью гидрокортизона. В опытах на кроликах и белых мышах определяли в динамике следующие иммунобиологические показатели: комплемент, пропердин, лизоцим, бактерицидность сыворотки крови, тканевую проницаемость, барьернофиксирующую функцию организма, незавершенный и завершенный фагоцитоз.
Однократное и курсовое (в течение 10 дней) введение кроликам всех исследуемых препаратов (пиридрол по 1 мг/кг, родозин и экстракт элеутерококка по 1 мл/кг) не сопровождалось существенными изменениями титра пропердина и лизоцимной активности сыворотки, пиридрол и родозин не влияли и на комплементарную активность, тогда как элеутерококк статистически значимо снижал титр комплемента. Все препараты при курсовом назначении уменьшали бактерицидную активность крови, отражающую функциональное состояние гуморальных факторов неспецифической иммунобиологической реактивности организма. Судя по результатам внутрикожной трипановой пробы Лещинского-Кавецкого, они повышали тканевую проницаемость, что может способствовать генерализации инфекционного процесса.
В опытах на белых мышах родозин (5 мл/кг), пиридрол (0,5 мг/кг) и экстракт элеутерококка (5 мл/кг) при однократном и курсовом введении угнетали барьернофиксирующую функцию организма. Это проявлялось повышенной распространяемостью листерий в организме зараженных животных, а также их большей смертностью. Препараты снижали количество активных нейтрофилов, их поглотительную и переваривающую способность. Нормализация описанных сдвигов реактивности организма происходила в течение 12—48 часов.
Таким образом, пиридрол, родозин и экстракт элеутерококка у интактных животных вызывают более или менее выраженное подавление неспецифической иммунобиологической реактивности организма, причем действие родозина и экстракта элеутерококка более выражено, чем пиридрола.
Учитывая роль патологического фона для выявления адаптогенного действия, следующая серия экспериментов была выполнена по аналогичному плану, но на фоне снижения иммунологической реактивности с помощью гидрокортизона. В соответствии с многочисленными литературными данными (Crepea и соавт., 1951; А. Н. Мешалова и И. Б. Фрезинова, 1960; 1 А. А. Триполитова, 1960; Н. И. Брауде и Е. В. Чернохвостова, я 1964 и др.) гидрокортизон (кроликам по 10 мг/кг внутримышечно в течение 3 дней, мышам — 125 мг/кг однократно) вызывал снижение титров комплемента и пропердина сыворотки, барьернофиксирующей функции и тканевой проницаемости, а также угнетение фагоцитарной активности лейкоцитов; титр лизоцима и бактерицидность сыворотки не изменились.
Для выяснения характера совместного действия гидрокортизона и психостимуляторов последние инъецировали кроликам в вышеуказанных дозах в течение 5 дней, затем животным вводили 3 дня гидрокортизон (10 мг/кг) совместно с психостимулятором и на протяжении последующих 15 дней только психостимулятор. Мыши получали гидрокортизон (125 мг/кг) однократно.
Снижение иммунологической реактивности гидрокортизоном на фоне профилактического введения пиридрола, родозина и экстракта элеутерококка не только не ослабевало, но, напротив, было более выраженным. Это проявлялось в более значительном снижении титров комплемента и пропердина, барьернофиксирующей функции и фагоцитарной активности лейкоцитов, увеличении тканевой проницаемости.
Таким образом, препараты родиолы и элеутерококка как, по-видимому, и другие представители группы женьшеня, не повышают неспецифическую иммунобиологическую реактивность организма. Очевидно, их не следует применять в коль плексе лечебных мероприятий при ряде инфекционных заболеваний ввиду возможного снижения сопротивляемости организма к инфекции.