Глава 17. Динамический бум в фитоценологии (Динамика растительности)
Растительное сообщество - явление очень динамичное. О некоторых чертах его динамики мы уже говорили, когда обсуждали вопросы разногодичной и сезонной изменчивости, постоянно подлаживающей сообщество к изменениям климата. Эти изменения обратимы: весна и осень случаются каждый год и даже более засушливые годы также рано или поздно повторяются, и потому, то состояние растительности, которое наиболее соответствовало этому сухому варианту климата (со снижением продуктивности, недоразвитием требовательных к влаге растений и т. д.), также рано или поздно обязательно повторится. Впрочем, естественно, уже усвоенный нами принцип индивидуальности видов и сообществ наводит на мысль, что, строго говоря, полного повторения не может быть и сообщество при обратимых изменениях каждый год в чем-то своеобразно. Тем не менее в годы со сходным климатом, при условии что предшествующие годы также отличались несильно, сообщество будет иметь примерно тот же вид, а этого при следовании парадигме континуума уже достаточно, чтобы отнести сообщество к одной и той же низшей классификационной единице.
Однако наряду с такими чисто обратимыми колебаниями состава и структуры сообщества вокруг какого-то среднего уровня существуют еще и другие изменения, которые носят названия необратимых. Создано несколько вариантов классификации таких изменений, но мы условимся делить их на два больших класса: эволюции и сукцессии. В первом случае в результате изменений растительности возникают совершенно новые сообщества в результате появления новых видов или из-за их миграции из других районов и формирования новых сочетаний популяций, во втором - формирования новых сочетаний популяций не происходит и вследствие процесса возникают сообщества, которые в данном районе уже имеются. Таким образом, сукцессии всегда в большей или меньшей степени - повторение пройденного.
Динамика растительности всегда была излюбленным вопросом фитоценологов, однако в последние 20 лет, соответствующие воцарению новой парадигмы, во всем мире произошло резкое возрастание интереса к вопросам динамики. И это отражало не столько новое видение растительности (хотя это тоже подталкивало фитоценологов к разработке вопросов теории динамики), сколько повышение плотности населения земного шара и резкое увеличение воздействия человека на биосферу. Ранее, когда человечество не было таким энерговооруженным и буферные свойства природы позволяли ей выдерживать давление антропогенного пресса, биосфера была подобна батуту, который, прогибаясь под давлением человека, был способен вернуться в прежнее состояние. Такие "прыжки на батуте" были безопасными вплоть до начала нынешнего столетия. Сегодня же положение меняется и озабоченное человечество говорит об экологическом кризисе, ищет пути его преодоления и потому, естественно, что вопросы о том, как меняются растительные сообщества под влиянием человека и как происходит восстановление растительности после нарушения, стали центральными для фитоценологов.
Увеличение народонаселения повлекло, с одной стороны, расширение площади пашни, а с другой - возрастание поголовья скота. Однако поскольку площадь земного шара при этом не изменилась и под пашню осваивались чаще земли, которые раньше использовались как пастбища, то их оставшаяся часть оказывалась как бы в тисках повышения пастбищной нагрузки из-за факторов сокращения площадей и одновременного возрастания поголовья пасущегося скота. Вырваться из этих тисков можно было только благодаря вовлечению в сельскохозяйственное использование лесов, занятые ими площади непосредственно превращались в пашню или становились лесными пастбищами, что вызывало их нарушение. Таким образом, перевыпас стал одним из самых главных факторов нарушения естественной растительности.
Однако увеличение плотности популяции человека на планете сопровождалось не только перевыпасом и сведением лесов, одновременно повышалась его энерговооруженность (а каждая новая тысяча калорий энергии - это новое загрязнение атмосферы, почвы, вод) и расширение использования средств химии для повышения урожая и борьбы с сорняками и вредителями.
Все эти факторы влияния человека на биосферу, взятые воедино, называются антропогенным прессом. Процессы в растительности, протекающие под его влиянием, и стали основным объектом исследований фитоценологов второй половины XX в., причем интерес их к этим процессам был столь велик, что можно даже говорить о своеобразном динамическом буме в фитоценологии.
Основным мерилом результативности ученого является объем печатной продукции. Было бы, конечно, чрезмерным оптимизмом считать любую научную статью истинным вкладом в науку (увы, создается немало и "научной макулатуры"), но все-таки по количеству научных публикаций, посвященных тому или иному вопросу, можно судить о том, насколько он привлекает внимание ученых.
Для того чтобы показать динамический бум в фитоценологии, я попросил своих сотрудников проанализировать динамику изменения публикаций по вопросам динамики растительности за последние десять лет по "Реферативному журналу". Это оригинальное научное издание, которое опирается на активность большого числа энтузиастов- референтов (к их числу относится и автор), публикует краткое содержание статей и книг, которые появляются в нашей стране и за рубежом. На рис. 20 приведена диаграмма, показывающая, насколько резко вырос интерес к вопросам динамики растительности, особенно антропогенной, за рубежом и в нашей стране. К примеру, общее число публикаций о динамике растительности в нашей стране возросло за этот период более чем в 10 раз, а число публикаций по антропогенной динамике - в 30 раз (!). За рубежом интерес к антропогенным изменениям в растительности пробудился несколько раньше, и потому при увеличении абсолютного количества работ о динамике растительности доля публикаций о влиянии антропогенного пресса сохранялась примерно такой же.
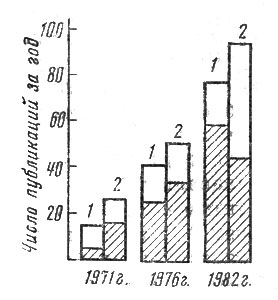
Рис. 20. "Валовая" печатная продукция отечественных (1) и зарубежных (2) фитоценологов, исследующих процессы динамики растительности. Заштрихованная часть столбиков - доля публикаций, посвященных вопросам антропогенной динамики
Однако о динамическом буме можно судить не только по таким формальным показателям. За последние годы вопросам динамики был посвящен ряд международных симпозиумов. В США переизданы заново фундаментальный труд Ф. Клементса о растительных сукцессиях и специальный факсимильный (т. е. воспроизводящий офсетным способом труды так, как они были изданы в свое время) выпуск наиболее значительных работ по этому вопросу, начиная от первых статей Ф. Клементса и вплоть до публикаций 70-х годов. Воззрения на сукцессии, как будет видно из дальнейших экскурсов в этот вопрос, у сторонников новой парадигмы вовсе не те, что были у организмистов. Тем не менее новая парадигма не может не впитывать все то, что дала старая, и потому подчеркнутое внимание к работам организмистов помогало разработке новой концепции. Самый авторитетный журнал фитоценологов - "Vegetatio" посвятил за эти годы проблемам динамики несколько выпусков, где публиковались работы континуалистов. В нашей стране появился целый ряд монографий, посвященных антропогенной динамике под влиянием осушения, сооружения гидротехнических комплексов на реках, изменению луговой растительности под влиянием удобрений и т. д.
Понятие "эволюция" применительно к растительности трактовалось организмистами в соответствии с аналогией эволюции растений. Считалось, что главным фактором эволюции является коадаптация, взаимоприспособление гидов, причем в целом сообщества развиваются под контролем все тех же "всемогущих" эдификаторов, и потому все сообщества одной формации представляют генетическое единство. Организмисты любили говорить о типах растительности-вкладчиках, которые объединяя свои виды (как растения свои гены), дают новый тип растительности. Наряду с эволюцией сообществ как целостных явлений подразумевалась и принципиальная возможность "гибридизации" типов растительности. Скажем, луга рассматривались как "гибрид" степей, альпийских высокотравий и сообществ лесных опушек.
Р. Уиттекер сформулировал иные представления об эволюции, которые как бы поставили точки над "и" в понимании растительности континуалистами. Эволюция носит сеткообразный характер, т. е. имеет место такое же индивидуальное распределение видов по градиенту эволюционного времени, как и вдоль экологических градиентов местообитаний. Таким образом, каждый вид приходит в сообщество своим путем и по-своему из него уходит, хотя, естественно, совместное сосуществование может датироваться многими тысячами лет. Но предпочтение патиентом какого-то виолента (скажем, пристрастие черники, кислицы и т. д. к обществу ели) не связано с какой-то внутренней фитоценотической связью, а определяется лишь тем режимом, который создает виолент. По этой причине, если несколько виолентов создают сходные режимы, то соответственно с этими режимами могут быть связаны одни и те же патиенты. Перечисленные виды встречаются и под пихтой, и даже под сосной, черника, кроме того, в условиях альпийской растительности вообще покидает полог деревьев и становится самостоятельным доминантом альпийских травяно-кустарниковых сообществ. На рис. 21 показаны две модели эволюции растительности - организмистская и континуальная.
![Рис. 21. Организмистская (коадаптационная) и континуальная (с дивергенцией ниш, сеткообразная) модели эволюции (по: [Уиттекер, 1980]), А-Д - типы сообществ](pic/000021.jpg)
Рис. 21. Организмистская (коадаптационная) и континуальная (с дивергенцией ниш, сеткообразная) модели эволюции (по: [Уиттекер, 1980]), А-Д - типы сообществ
В первом случае виды коэволюционируют комплектами, и в итоге следует ожидать формирования дискретных сообществ. Во втором - все виды эволюционируют индивидуально, часть их в ходе эволюции теряется и замещается вновь появляющимися вследствие миграции или видообразования видами. Из двух нечетко выраженных типов сообществ вследствие эволюции возникло три столь же нечетко различимых сообщества.
Природная эволюция - изменения растительности геологического масштаба времени, десятков и сотен тысяч лет, однако последние столетия проявился новый процесс, который называют по-разному - синантропизацией, антропогенизацией и т. д., но мне кажется, что называть его предпочтительнее антропогенной эволюцией. При этом происходят и появление новых видов (скажем, выведение новых сортов культурных растений, отличия которых от ранее существовавших нередко имеют масштаб вида), и целенаправленное комбинирование видов в новые сочетания (разные типы лесных посадок, сеяных лугов, пашенных сообществ), и сопутствующие деятельности человека гигантские стихийные перестройки растительности в результате миграции многих видов из одного района в другой и из-за уничтожения многих компонентов естественных сообществ. Эволюция не предполагает обязательный прогресс (скажем, формирование паразитных червей с упрощением их организмов из-за перехода на харчи хозяина, как, впрочем, и утеря хлорофилла и сокращение поверхности у паразитных растений - заразихи и повилики, - это также эволюция). По этой причине, несмотря на всю нежелательность таких изменений, сопровождающихся выпадением видов (недаром сейчас существует целая армия ботаников, занимающаяся проблемой мониторинга за редкими и исчезающими видами!), эти изменения также вполне правомерно подвести под широкую рубрику антропогенной эволюции. В этом случае также изменения протекают сеткообразно, и как результат происходит переупаковка ниш, хотя если при прогрессивной эволюции ниши все более сужаются и происходит "втискивание" новых популяций и даже порой "выталкивание" более слабых, не обладающих достаточным "чувством коллективизма", то в этом случае ниши видов, наиболее адаптированных к антропогенному воздействию, расширяются.
Примеров антропогенной эволюции приведено в фитоценологической литературе великое множество. Так, появление европейцев и их черных спутников-рабов в Южной Америке вызвало процесс, который в научной литературе получил название "африканизация американских саванн". Крупные стада, которые появились вместе с европейцами, вызвали разрушение травяного яруса саванн. И в эти нарушенные местообитания на правах R-стратегов-эксплерентов стали внедряться африканские виды злаков, семена которых попали в Америку вместе с черными рабами. В Австралии одичала и широко распространилась опунция (кактус), которую завезли туда для создания колючих изгородей.
Новые и неведомые ранее человеку сообщества появились на техногенных субстратах, химический состав которых в корне отличался от свойств покрывающих планету почв. В водоемах Европы появились новые сообщества, где европейские виды сообитают совместно с элодеей канадской и т. д.
Антропогенной эволюцией можно считать и создание новых типов полевых и лесных сообществ, но об этом - в заключающей книгу главе.
Что касается прогрессивной антропогенной эволюции, то ее достижения не менее значительны и иллюстрируются любым фактом интродукции новых видов. Над решением этой проблемы трудятся многочисленные коллективы ботанических садов, и если, скажем, взглянуть на немалые площади побережья Крыма, где вместе с видами местной флоры произрастают многие американские виды, подобные секвойядендронам, средиземноморские кипарисы, пальмы из Океании и десятки других "гостей", то в целом весь этот сложный комплекс видов должен рассматриваться как результат целенаправленной эволюции растительности под влиянием человека.
В отличие от "новаторств эволюции" сукцессии повторяют уже пройденные пути, и если после зарастания болота формируется лес, то такой лес уже где-то поблизости существует, как существуют сообщества скал или тополевые леса, развившиеся на новых отложениях песка вдоль русла.
Интерес к сукцессиям в период становления органимистской парадигмы был очень велик, и наиболее полно выразил организмистские представления о сукцессии выдающийся американский ученый Ф. Клементс. То, что среди трех фотографий фитоценологов в последнем издании книги Т. А. Работнова ("Фитоценология") помещен портрет Ф. Клементса (вместе с ним помещены портреты Ж. Браун-Бланке и В. Н. Сукачева), не случайно и вполне заслуженно, так как без поиска "философского камня" не возникли бы современные представления о природе сукцессий.
По своему характеру Ф. Клементс был дедуктивист с железной логикой. Он сформулировал представления о сукцессии, которая протекает под влиянием чисто внутренних факторов сообщества, которое меняется (в целом в соответствии со вторым законом термодинамики, хотя это скажут последователи Клементса уже после его смерти) в направлении достижения состояния наибольшего соответствия климату. Таким образом, по Клементсу, в районе с более или менее однородным климатом все сообщества должны рано или поздно прийти к одному и тому же состоянию, преобразовав местообитания и сделав их также равновесными с климатом. Озера должны зарасти и высохнуть, пески - оглиниться, камни - выветриться, сухие местообитания - стать более увлажненными и т. д. Если климат соответствует, скажем, тайге, то везде сформируются сообщества ельников, если степи - разнотравно-ковыльная степь, пустыне - сообщества ксерофильных кустарников. В какой-то степени районы климакса Ф. Клементса похожи на зонобиомы Г. Вальтера, хотя в отличие от Клементса Вальтер допускает в пределах своего зонобиома и сообщества, которые значительно отличаются от преобладающих зональных (например, болота, сообщества солончаков).
Эта концепция заключительного сообщества получила название концепции моноклимакса. Ее заслуженно критиковали многие ученые с самого момента появления, но Ф. Клементс был увлечен этой идеей и преданно пронес ее, как знамя, через всю свою научную биографию. К тому времени, как в самом начале 50-х годов на фитоценологическом горизонте вспыхнула ослепительная звезда Р. Уиттекера, Ф. Клементс уже ушел из жизни, хотя в последней работе вновь подтвердил свою верность идее моноклимакса и назвал ее ""Сезам, открой дверь" для фитоценологии". Р. Уиттекер очень остроумно дополнил эту фразу словами "в пустую комнату". Развивая сформулированное оппонентами Ф. Клементса положение о поликлимаксе (принципиальной возможности формирования в одном районе нескольких устойчивых сообществ, соответствующих данному климату, но связанных с разными субстратами), Уиттекер сформулировал положение о климаксе-континууме, т. е. гамме постепенных переходов между разными климаксовыми сообществами.
Однако, если концепция моноклимакса была раскритикована уже в 50-х годах, серьезные возвраты к ней периодически случаются. Так, в 1981 г. была опубликована монография С. М. Разумовского о концепции моноклимакса и дискретности в растительности. Кстати, С. М. Разумовский был также блестящим дедуктивистом, способным к выстраиванию безупречных в отношении логики последовательностей представлений, но с неверной первой посылкой.
Тем не менее в теоретическом наследии Клементса наряду с концепцией моноклимакса были еще и представления о том, как протекают сукцессии, т. е. о "механике сукцессий". Очередь критики этих представлений настала лишь в 70-х годах, когда на них обрушился шквал критики континуалистов.
Как виделась сукцессия Ф. Клементсу? Он считал, что если сообщество неравновесно по отношению к климату (сформировалось на скалах, в озере и т. д.), то оно будет переделывать среду до тех пор, пока не достигнет уровня максимальной для данной зоны продуктивности и наиболее развитой структуры. Под сообществом возникает почва с развитым профилем (подзол, чернозем и т. д.). При этом будут происходить процессы конвергенции сообществ разных исходных местообитаний и климаксовое сообщество станет превалирующим по площади. Виды при этом как бы разбиваются на четкие группы, которые постепенно сменяют друг друга вследствие того, что их предшественники настолько изменили среду, что она стала неблагоприятной для них, но гостеприимным кровом для видов последующей группы. Классическим примером тому было зарастание скал: группа накипных лишайников, после того как в достаточной мере выветрит скалу, сменялась кустовыми лишайниками и мхами. Те активно брались за дело и продолжали выветривание до тех пор, пока слой мелкозема не позволял поселиться травам. Далее следовали кустарники и затем - деревья. Так же мыслился процесс зарастания озера, когда четко сменялись группы плавающих растений (рдест и пр.), потом - прикрепленных ко дну кувшинок, лилий и т. д., потом - полупогруженных растений - рогоза, тростника, камыша и т. д., далее - осок. После этого озеро превращалось в низинное травяное болото, и, по мере того как слой торфа увеличивался, он обеднялся минеральными элементами (связь с минеральным дном уже была прервана, так как корни растений не достигают дна и, питаясь за счет торфа, растения не могут извлечь все накопленные в торфе минеральные элементы), на смену такому низинному болоту приходило вначале переходное, а затем и верховое, со сфагновыми мхами и сосной. В свою очередь, верховое болото могло постепенно перейти в климакс таежной зоны - еловый лес.
Эта безупречная в отношении логики схема никогда и никем не была подкреплена фактическим материалом. Основным методом изучения сукцессий был дедуктивный. Последовательность сосуществующих в пространстве сообществ интерпретировалась как временной ряд, который никто не оспаривал, но и никто не проверял. Просто к этому привыкли и считали, что иначе не может быть! Если кто-то видел ряд поясов на озере - от центра к берегу как раз и расположены последовательно те группы растений, о которых только что говорилось, - то он считал, что по мере зарастания в результате отложений органического вещества "трупов" растений озеро будет мелеть и все пояса начнут постепенно смещаться к центру, причем в обязательной и строгой последовательности. Когда виды группы А сменяют виды группы В, виды группы С - виды группы В, виды группы D - виды группы С и т. д., и при этом смена будет происходить потому, что эффекты влияния на среду предшествующей группы всегда полезны для той, что идет за ней по пятам.
Р. Уиттекер смог увидеть, что в логической конструкции Клементса неверен сам фундамент - организмистские представления о целостности и жесткой детерминированности сообществ и последовательности их смены во времени. Впрочем, начав с язвительной полемики с Клементсом, Р. Уиттекер в последних работах давал ему высокую оценку и уже упомянутое переиздание наиболее фундаментальной научной монографии Ф. Клементса "Сукцессии и растения-индикаторы" также является данью уважения этому неутомимому искателю "философского камня".
В 70-е годы представления о климаксе-континууме уже стали привычными и настала очередь критики второго фундаментального положения концепции Ф. Клементса - жесткой детерминированности сукцессии. В это время, как образно заметил один из видных современных фитоценологов Дж. Майлс, "изучение сукцессий перешло от стадии догматизма к прагматизму". В данном случае не следует рассматривать прагматизм в чисто философском плане, Майлс имел в виду в первую очередь реалистичность подхода, стремление обосновать высказываемые точки зрения фактами. В основу анализа сукцессий в это время были положены не косвенные методы пространственных экстраполяций во временные ряды, на основании которых, к примеру, была создана схема зарастания озер, а прямые наблюдения за изменениями сообществ, сюда входили анализ геоботанических карт, составленных в разные годы, ботанический анализ торфов, хранящих историю смены тех растений из "трупов", которые формировали его разные слои, спорово-пыльцевой анализ, позволяющий по наборам спор, накапливающихся, скажем, в отложениях на дне озера, выявить последовательность смены видов в сукцессиях.
В результате использования этих прямых методов были накоплены данные, показавшие, что модель Клементса верна не всегда.
Во-первых, было установлено, что последовательность видов может варьировать в достаточно широких пределах. Дело дошло до того, что для описания сукцессий стали применять широко распространенную в математике модель марковских цепей, т. е. такой последовательности событий, когда взаимообусловленность всех звеньев ряда А-B-C-D-E и т. д. очень низкая и В зависит только от А, С - от В, D - от С и т. д.
Во-вторых, оказалось, что случаи "патриотичности" поведения видов в сукцессии также весьма частные и далеко не всегда предшественники так "заботятся" о своих последователях. Сплошь и рядом отношения могут носить обратный характер и предшественники "не пускают" виды следующей стадии, которые водворяются только в том случае, если, скажем, столь негостеприимных предшественников потопчут ногами олени и создадут необходимые "дырки" в сообществе. Так бывает, к примеру, на пустошах, где после гарей разрастаются мхи, через которые не могут пробиться вересковые кустарнички. Кроме того, иногда виды последующих фаз могут внедриться лишь потому, что они оказываются более и более устойчивыми к неблагоприятному режиму предшественников. Вначале селятся деревья-солнцелюбы, а под их полог постепенно "набиваются" все более и более тенелюбивые патиенты. Руководствуясь этим принципом, ставропольский ботаник В. В. Скрипчинский смог воспроизвести близкие к естественным участки лесов.
Наконец, возможен случай, когда предшественники вообще не влияют на поселенцев и смена происходит просто вследствие того, что у этих видов разные стратегии. Первые поселенцы могут представлять растения-эксплеренты с банком семян и слабо выраженным свойством К, а последующие виды, семена которых притащили муравьи или принесли птицы. Эти виды имеют более развитое свойство К и потому, протиснувшись в сообщество, занимают ниши, изгоняя из него эксплерентов.
Работа американских фитоценологов Коннела и Слайтера, которая была опубликована в журнале "Американский натуралист" в 1977 г. и содержит описание различных моделей сукцессий, так понравилась континуалистам, что стала, пожалуй, на сегодня своего рода рекордсменом цитируемости.
Таким образом, ревизия воззрений Клементса коснулась и понимания климакса, и трактовки механизма сукцессий, причем его оппоненты усиленно экспериментировали (особенно часто исследовались сукцессии зарастания заброшенной пашни, вспомним и описанные в главе о стратегиях остроумные эксперименты Периша и Безаса). Кроме того, сам центр тяжести интересов фитоценологов-континуалистов сместился с исследования первичных сукцессий, которые были объектом для умозаключений Ф. Клементса, на вторичные антропогенные сукцессии, которые были связаны с деятельностью человека.
Среди антропогенных сукцессий различают две большие группы. Первая группа - это изменения, которые протекают под непосредственным воздействием одного из антропогенных факторов - постоянного выпаса, осушения, орошения, удобрения, действия атмосферных загрязнителей, радиации и т. д. В этом случае изменения могут сопровождаться повышением продуктивности (если внешний фактор увеличивает гиперпространство ниш), как при удобрении или орошении, или, напротив, снижать ее. В последнем случае процесс носит название ретрогрессии. Классическим примером ретрогрессии является изменение состава растительности под влиянием выпаса. В степях, где при сенокосном использовании господствуют ковыли и разнотравье, в этом случае вначале доминирование переходит к типчаку, который значительно устойчивее ко всему комплексу факторов выпаса: уплотнению почвы, вытаптыванию и постоянному скусыванию побегов. Однако если интенсивность выпаса еще выше, то не выдерживает и типчак и массово развивается полынь австрийская, которая в условиях степной зоны является своеобразным чемпионом устойчивости к выпасу. При внесении удобрений чемпионами могут быть одуванчик лекарственный, виды подорожника, клевер ползучий и "чемпион чемпионов" спорыш, или птичья гречишка. Примерно так же протекают сукцессии под влиянием рекреационных нагрузок, т. е. туризма, выездов за город и т. д. В травостое лесов происходит постепенная замена типичных лесных видов пастбищными луговыми, исчезает возобновление древесных пород, и потому древостой со временем становится более редким.
Очень интересны сукцессии под влиянием радиации, которые описали Р. Уиттекер и Дж. Вудвелл в эксперименте на о-ве Лонг. Оказалось, что по мере увеличения дозы радиации сукцессия оказывается как бы зеркальным отражением той, что протекает при зарастании скал, и это было положено Ф. Клементсом в основу его общих представлений о данном процессе. Вначале исчезают деревья, потом кустарники, мхи и лишайники, и, наконец, наиболее устойчивыми к действию радиации оказываются сохраняющиеся в почве одноклеточные водоросли. В такой же последовательности, кстати, происходят изменения состава и структуры сообществ и при сильном загрязнении например при осаждении пыли некоторых предприятий горнодобывающей промышленности и при "переудобрении" - на территории вокруг предприятий, производящих минеральные удобрения.
Однако, если действие вызвавшего сукцессию фактора прекратилось, начинается обратный процесс восстановления. Причем восстановительный процесс протекает уже, как говорят фитоценологи, чисто автогепетически, т. е. аналогично первичной сукцессии Ф. Клементса по причине, заложенной в самом сообществе и экосистеме, которые "стремятся" прийти в соответствие с тем комплексом факторов среды, который восстанавливался после действия антропогенного фактора.
На пастбище начинается процесс постпастбищной демутации - постепенно вновь появляются (или из-за сохранившихся в почве подземных органов вегетативного размножения "банка семян", или из-за семян, доставляемых животными и ветром) те же виды, которые характерны для сенокосной степи. В лесу после сведения древостоя, после рубки или пожара начинается процесс восстановления, причем вначале буйно развиваются травы, которые получили дополнительные ресурсы в результате исчезновения полога деревьев, разложения их корней или минеральных веществ, вымываемых в почву из золы. Далее могут восстановиться те породы, которые были уничтожены, или появляется "буферная" порода, как, например, ольха в ельниках. И уже под пологом обладающей свойством R ольхи начинается восстановление ели, которая из-за свойства S хорошо чувствует себя под сенью полога ольхи и затем, когда растения вырастут, за счет свойств К подавляет ее.
Полное восстановление возможно не во всех случаях, и потому после сильных воздействий человека экосистема может не приходить к прежнему состоянию. На границе лесов и степи сведение леса способствует формированию уже не лесной, а степной растительности. После пожара и выпаса на месте тропического леса может формироваться саванна. После забрасывания пашни залежи многие десятки лет остаются отличными по видовому составу от исходной степи или прерии. Плохо восстанавливаются сообщества тундры после выпаса крупных стад оленей; наконец, перевыпас в горах может вызвать развитие процессов эрозии, и в этом случае после потери мелкоземистого слоя восстановление затягивается на тысячелетия.
Иными словами, далеко не после всякого воздействия человека "экологический батут" может восстановить растительность. В ряде случаев человеку приходится помогать ликвидации последствий собственного (часто неосмотрительного) воздействия на природу - проводить посадку леса, высевать травосмеси. Особой заботой для человечества является восстановление растительности на промышленных техногенных субстратах - отвалах дорог, намытых грунтах, выбросах пустой породы из шахт и т. д. Площадь их непрерывно увеличивается, и потому превратить их в полезные экосистемы, дающие кислород и растительную массу,- задача весьма актуальная.
В ряде случаев, если субстрат более или менее благоприятен, природа справляется сама и возникновение сомкнутой растительности происходит чисто автогенетически, причем постепенно в такой породе формируется зачаточный гумусовый слой и сукцессия идет по клементовскому типу стимуляции. Однако нередко такой сукцессии помогают - покрывают породу слоем чернозема и высевают травы или высаживают деревья. В США покрывают отвалы слоем лесной почвы, которая содержит банк семян, и тогда происходит быстрое и успешное восстановление древесной растительности. Оригинальный быстрый способ заселения нарушенных участков предложил советский фитоценолог Д. С. Дзыбов. Он покрывает такой участок слоем мелкого высушенного и нарезанного, скошенного в три срока степного сена. В результате семена сразу десятков видов трав попадают в почву в тесном естественном комплекте и за 2-3 года восстанавливается травостой, состав которого очень близок к истинной степи. В Прибалтике на терриконах пустой породы, образующихся при добыче сланцев, после покрытия их слоем плодородной почвы и посадки деревьев создают скверы и парки.
Изучение антропогенных смен растительности - дело очень интересное. Объектов для исследования много, смены протекают достаточно быстро, и несложно измерить время действия фактора и его интенсивность. Тем не менее не это является движителем динамической проблематики в фитоценологии, которая привела к "буму". Все эти исследования нацелены на организацию мониторинга состояния растительности, разработку мер расчета устойчивости разных типов сообществ к разным воздействиям человека, разработку способов стимулирования восстановительной сукцессии и подтормаживания нарушающих биосферу аллогенных смен. Если таких исследований не вести и если практикам не учитывать их результатов и рекомендаций науки, батут может не выдержать... и среда существования человечества уподобится тому варианту модели сукцессии, который называется ретрогрессией. Нашим потомкам придется жить хуже, чем нам. Но этого не должно случиться.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'